Mephedron (4-methylmethcathinone) er et β-ketoamfetamin-stimulerende misbrugsstof med tætte strukturelle og mekanistiske ligheder med metamfetamin. En af de mest kraftfulde virkninger forbundet med mephedron er evnen til at stimulere frigivelse af dopamin (DA) og blokere dets genoptagelse gennem dets interaktion med dopamintransportøren (DAT). Selvom mephedron ikke forårsager toksicitet for DA-nerveender, kan dets evne til at fungere som en DAT-blokker give beskyttelse mod metamfetamininduceret neurotoksicitet ligesom andre DAT-hæmmere. For at teste denne mulighed blev mus behandlet med mephedron (10, 20 eller 40 mg/kg) før hver injektion af et neurotoksisk regime af metamfetamin (4 injektioner af 2,5 eller 5,0 mg/kg med 2 timers mellemrum). Integriteten af DA-nerveender i striatum blev vurderet gennem målinger af DA-, DAT- og tyrosinhydroxylase-niveauer. Den moderate til alvorlige DA-toksicitet, der var forbundet med de forskellige doser af metamfetamin, blev ikke forhindret af nogen dosis af mephedron, men blev faktisk væsentligt forstærket. Hypertermi forårsaget af kombineret behandling med mephedron og metamfetamin var den samme som set efter et af stofferne alene. Mephedron forstærkede også de neurotoksiske virkninger af amfetamin og MDMA på DA-nerveender. I modsætning hertil beskyttede nomifensin mod metamfetamin-induceret neurotoksicitet. Da mefedron øger metamfetamins neurotoksicitet, tyder de nuværende resultater på, at det interagerer med DAT på en måde, der er forskellig fra andre typiske DAT-hæmmere. De relativt uskadelige virkninger af mephedron alene på DA-nerveender maskerer en potentielt farlig interaktion med stoffer, der ofte misbruges sammen med det, hvilket fører til øget neurotoksicitet.
Mephedron (4-methylmethcathinon) er et cathinonderivat og en strukturel analog til metamfetamin og 3,4-methylendioxy-metamfetamin (MDMA). Mefedron er en psykoaktiv ingrediens i "badesalte" sammen med andre forbindelser som methylon, butylon og 3,4-methylendioxypyrovaleron (MDPV). β-ketoamfetaminer misbruges i stigende grad, hvilket ikke mindst skyldes den meget begrænsede tilgængelighed af prækursorer, der er nødvendige for at syntetisere metamfetamin og MDMA i hemmelige laboratorier, og en tilsvarende reduktion af deres renhed (Winstock et al. 2011b, Brunt et al. 2011). I takt med at misbruget af β-ketoamfetamin fortsætter med at stige, er listen over skadevirkninger vokset til at omfatte kardiovaskulære komplikationer, uro, søvnløshed, psykose og depression (Schifano et al. 2011, Prosser og Nelson 2012).
Som kemisk beslægtet med metamfetamin og MDMA er det ikke overraskende, at β-ketoamfetaminerne har mange af de samme virkninger på centralnervesystemet som de førstnævnte stoffer. For eksempel blokerer disse stoffer dopamin- (DA) og serotonin- (5-HT) transportører (henholdsvis DAT og SERT) (Cozzi et al. 1999, Rothman et al. 2003, Fleckenstein et al. 2000, Lopez-Arnau et al. 2012), og de stimulerer frigivelse af monoaminer in vitro (Kalix og Glennon 1986, Gygi et al. 1997, Rothman et al. 2003) og in vivo (Gygi et al. 1997, Kehr et al. 2011). Methcathinone forårsager vedvarende reduktioner af tryptophan hydroxylase og tyrosin hydroxylase (TH) aktivitet og udtømning af DA og 5-HT (Gygi et al. 1997, Gygi et al. 1996, Sparago et al. 1996). PET-billeddannelsesundersøgelser hos afholdende methcathinonbrugere afslørede reduceret striatal DAT-tæthed, hvilket tyder på et tab af DA-terminaler (McCann et al. 1998). Den samtidige stimulering af DA-frigivelse og hæmning af optagelsen afspejler de kritiske elementer, der ligger til grund for den neurotoksicitet, der er forbundet med metamfetamin (Kuhn et al. 2008, Yamamoto og Bankson 2005, Cadet et al. 2007, Fleckenstein et al. 2007).
Vi (Angoa-Perez et al. 2012) og andre (Baumann et al. 2012, Hadlock et al. 2011) undersøgte for nylig muligheden for, at mephedron kunne forårsage neurotoksicitet ligesom metamfetamin og MDMA. Overraskende nok var mephedron ikke giftigt for DA-nerveender i striatum (Hadlock et al. 2011, Baumann et al. 2012, Angoa-Perez et al. 2012). Spørgsmålet om, hvorvidt mephedron skader 5-HT-nerveender, er stadig uafklaret, da en undersøgelse dokumenterede positive effekter (Hadlock et al. 2011), mens en anden var negativ (Baumann et al. 2012). I lyset af den relativt godartede effekt af mephedron på DA-nerveender og i betragtning af dets egenskaber som DAT-blokker, antog vi, at det faktisk kunne beskytte DA-neuronsystemet mod de neurotoksiske virkninger af metamfetamin, ligesom det vides at forekomme med andre DAT-blokkere såsom amfonsyre (Pu et al. 1994, Schmidt og Gibb 1985, Marek et al. 1990) og nomifensin (Poth et al. 2012). Vi rapporterer nu, at mephedron øger metamfetamins neurotoksicitet betydeligt. Denne effekt gælder også for amfetamin og MDMA, stoffer, der ofte misbruges sammen med mephedron (Feyissa og Kelly 2008, Schifano et al. 2011). Disse overraskende resultater sætter misbruget af mephedron i et nyt lys og gør det endnu mere presserende at få øjnene op for denne subtile og farlige egenskab ved β-ketoamfetamin.
Materialer og metoder
Stoffer og reagenser
Mephedronhydrochlorid og 3,4-methylendioxymetamfetamin (MDMA) blev anskaffet fra NIDA Research Resources Drug Supply Program. (+) Metamfetaminhydrochlorid, nomifensinmaleat, d-amfetaminsulfat, pentobarbital, DA og alle buffere og HPLC-reagenser blev købt fra Sigma-Aldrich (St. Louis, MO, USA). Bicinchoninic acid protein assay kits blev købt hos Pierce (Rockford, IL, USA). Polyklonale antistoffer mod TH fra rotter blev fremstillet som tidligere beskrevet (Kuhn og Billingsley 1987). Monoklonale antistoffer mod rotte-DAT blev generøst leveret af Dr. Roxanne A. Vaughan (University of North Dakota, Grand Forks, ND, USA). HRP-konjugerede anti-IgG sekundære antistoffer blev leveret af Jackson ImmunoResearch Laboratories, Inc. (West Grove, PA, USA).
Dyrene
C57BL/6-hunnemus (Harlan, Indianapolis, IN, USA), der vejede 20-25 g på forsøgstidspunktet, blev anbragt 5 pr. bur i store skotøjsbure i et lyst (12 timer lys/mørk) og temperaturkontrolleret rum. Der blev brugt hunmus, fordi de er kendt for at være meget følsomme over for neuronale skader af de neurotoksiske amfetaminer og for at opretholde konsistensen med vores tidligere undersøgelser af metamfetamins neurotoksicitet (Thomas et al. 2010, Thomas et al. 2008, Thomas et al. 2009). Musene havde fri adgang til mad og vand. Udvalget for institutionel pleje og brug af Wayne State University godkendte dyreplejen og de eksperimentelle procedurer. Alle procedurer var også i overensstemmelse med NIH Guide for the Care and Use of Laboratory Animals.
Farmakologiske, fysiologiske og adfærdsmæssige procedurer
Mus blev behandlet med mephedron ved hjælp af et binge-lignende regime bestående af 4 injektioner på 10, 20 eller 40 mg / kg med et 2 timers interval mellem hver injektion. Denne binge-behandling, når den bruges til at injicere substituerede amfetaminer og katinonderivater, resulterer i omfattende DA-nerveskader. De doser af mephedron, der anvendes i øjeblikket, har tidligere vist sig ikke at være giftige for DA-nerveender (Angoa-Perez et al. 2012). Mus blev behandlet med metamfetamin (4X 2,5 eller 5 mg/kg), amfetamin (4X 5 mg/kg) eller MDMA (4X 20 mg/kg) alene eller i kombination med mephedron. Når musene blev behandlet med to stoffer, fik de en mephedron-injektion 30 minutter før hver af de fire injektioner af metamfetamin, amfetamin eller MDMA. Kontrolgruppen fik indsprøjtninger med fysiologisk saltvand på samme tidspunkt som for mephedron alene eller i kombination med andre amfetaminer. Som kontrol for virkningerne af en DAT-hæmmer på metamfetamintoksicitet blev musene behandlet med nomifensin (4X 5 mg/kg) 30 minutter før hver injektion af metamfetamin (4X 5 mg/kg). Alle indsprøjtninger blev givet via i.p.-ruten. Musene blev ofret 2 dage efter den sidste lægemiddelbehandling, når den amfetamin-associerede neurotoksicitet havde nået sit maksimum. Kropstemperaturen blev overvåget ved hjælp af telemetri med IPTT-300 implanterbare temperaturtranspondere fra Bio Medic Data Systems, Inc. (Seaford, DE, USA). Temperaturerne blev registreret ikke-invasivt hvert 20. minut, startende 60 minutter før den første METH-injektion og fortsatte i 9 timer derefter ved hjælp af DAS-5001-konsolsystemet fra Bio Medic.
Bestemmelse af striatal DA-indhold
Striatalvæv blev dissekeret bilateralt fra hjernen efter behandling og opbevaret ved -80 ° C. Frosset væv blev vejet og sonikeret i 10 volumener 0,16 N perchlorsyre ved 4 °C. Uopløseligt protein blev fjernet ved centrifugering, og DA blev bestemt ved HPLC med elektrokemisk detektion som tidligere beskrevet for metamfetamin (Thomas et al. 2010, Thomas et al, 2009).
Bestemmelse af TH- og DAT-proteinniveauer ved immunoblotting
Virkningerne af lægemiddelbehandlinger på striatal TH- og DAT-niveauer blev bestemt ved immunoblotting som et indeks for toksicitet for striatal DA-nerveender. Mus blev ofret ved halshugning efter behandling, og striatum blev dissekeret bilateralt. Vævet blev opbevaret ved -80 °C. Frosset væv blev opløst ved sonikering i 1 % SDS ved 95 °C, og uopløseligt materiale blev sedimenteret ved centrifugering. Protein blev bestemt ved hjælp af bicinchoninsyremetoden, og lige store mængder protein (70 μg/lane) blev opløst ved SDS-polyacrylamidgelelektroforese og derefter elektroblottet til nitrocellulose. Blots blev blokeret i Tris buffered saline indeholdende Tween 20 (0,1% v/v) og 5% fedtfri tørmælk i 1 time ved stuetemperatur. Primære antistoffer mod TH (1:1000) eller DAT (1:1000) blev tilsat til blots og fik lov til at inkubere i 16 timer ved 4 °C. Blots blev vasket 3 gange i Tris-bufferet saltvand for at fjerne ureagerede antistoffer og blev derefter inkuberet med HRP-konjugeret anti-IgG sekundært antistof (1:4000) i 1 time ved stuetemperatur. Immunreaktive bånd blev visualiseret ved forbedret kemiluminescens, og de relative tætheder af TH- og DAT-reaktive bånd blev bestemt ved billeddannelse med en Kodak Image Station (Carestream Molecular Systems, Rochester, NY, USA) og kvantificeret ved hjælp af ImageJ-software (NIH).
Analyse af data
To-vejs ANOVA'er blev udført for at analysere dosisvirkningerne af methamphetamin versus mephedron på DA, DAT og TH. Virkningerne af lægemiddelbehandlinger på striatal DA, TH og DAT-indhold blev testet for signifikans ved envejs ANOVA efterfulgt af Tukey's multiple sammenligningstest. Resultater af lægemiddelbehandlinger på kernekropstemperatur over tid blev analyseret ved hjælp af en tovejs ANOVA efterfulgt af Bonferronis test for at bestemme betydningen af forskelle i temperatur på individuelle tidspunkter efter behandling. Forskelle blev betragtet som signifikante, hvis p < 0,05. Alle statistiske analyser blev udført ved hjælp af GraphPad Prism version 5.02 til Windows (GraphPad Software, San Diego, CA, USA, www.graphpad.com).
Gå til:
Resultater
Virkninger af mephedron på metamfetamin-induceret neurotoksicitet
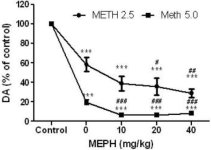
Mephedron i doser (10, 20 eller 40 mg/kg), der vides ikke at forårsage toksicitet i DA-nerven (Angoa-Perez et al. 2012), blev administreret 30 minutter før hver injektion af metamfetamin. Metamfetamin blev givet i doser, der forårsager moderat (4X 2,5 mg/kg) eller alvorlig (4X 5 mg/kg) skade på DA-nerveender i striatum (Thomas et al. 2004, Thomas et al. 2010). Resultater præsenteret i fig. 1 viser, at hovedeffekterne af metamfetamindosis (F1,40 = 66,60, p < 0,0001) og mephedron-dosis (F4,40 = 131,3, p < 0,0001) på DA-niveauer i striatum var meget signifikante ved tovejs ANOVA. Hovedeffekten af mephedron givet i kombination med enten 2.5 mg / kg (F4,22 = 35.96, p <0.001) eller 5.0 mg / kg methamphetamin (F4,17 = 953.9, p <0.0001) var også meget signifikant ved envejs ANOVA. Alle behandlinger med enten dosis metamfetamin ± mephedron forårsagede signifikant større reduktioner i DA sammenlignet med den respektive kontrol (p <0.0001 for alle). Fig. 1 viser også, at mephedron-doser på 20 (p < 0,01) og 40 mg/kg (p < 0,001) signifikant forstærkede de nedbrydende virkninger af 2,5 mg/kg metamfetamin på DA, mens alle doser af mephedron signifikant forstærkede virkningerne af 5,0 mg/kg metamfetamin på DA-niveauer (p < 0,0001 for alle).
Fig.1
Virkninger af mephedron på metamfetamininducerede reduktioner i striatal DA. Mus blev behandlet med de angivne doser af mephedron (MEPH) 30 minutter før hver injektion af 2.5 (-) eller 5.0 mg / kg (■) methamphetamin (METH) og ofret 2d senere til bestemmelse af striatal niveauer af DA ved HPLC. Data er gennemsnit ± SEM for 5-7 mus pr. Gruppe. Nogle fejlbjælker var for små til at overstige størrelsen på symbolerne og vises ikke synlige. ***p < 0,001 vs kontroller og #p < 0,01, ##p < 0,001 eller ###p < 0,0001 vs den respektive dosis metamfetamin (Tukey's multiple sammenligningstest).
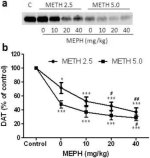
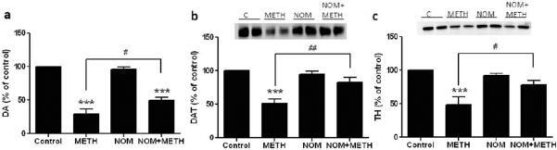
Fig. 2a viser, at mephedron signifikant øgede metamfetamininducerede reduktioner i DAT-niveauer som bestemt ved immunoblotting. Immunoblots blev kvantificeret, og i overensstemmelse med resultaterne for DA var hovedeffekterne af metamfetamindosis (F1,92 = 9.48, p <0.001) og mephedronedosis (F4,92 = 37.56, p <0.0001) på DAT-niveauer i striatum meget signifikante ved tovejs ANOVA (Fig. 2b). Hovedeffekten af mephedron givet i kombination med enten 2.5 mg / kg (F4,56 = 15.55, p <0.0001) eller 5.0 mg / kg metamfetamin (F4,39 = 24.84, p <0.0001) var også meget signifikant ved envejs ANOVA. Alle behandlinger med enten dosis metamfetamin ± mephedron forårsagede signifikant større reduktioner i DAT sammenlignet med den respektive kontrol (p <0.01 for 2.5 mg / kg metamfetamin alene; p <0.0001 for alle andre behandlinger). Fig. 2b viser også, at mephedron-doser på 20 mg / kg (p <0.01) og 40 mg / kg (p <0.001) signifikant forbedrede reduktionerne i DAT forårsaget af 2.5 mg / kg methamphetamin, mens kun 40 mg / kg mephedron-dosis signifikant forbedrede (p <0.01) virkningerne af 5.0 mg / kg methamphetamin på DAT-reduktioner.
Fig.2
Effekter af mephedron på metamfetamin-inducerede reduktioner i striatal DAT. Mus blev behandlet med de angivne doser af mephedron (MEPH) 30 minutter før hver injektion af 2.5 (●) eller 5.0 mg / kg (■) metamfetamin (METH) og ofret 2d senere til bestemmelse af striatal niveauer af DAT ved immunoblotting (a). Blots blev kvantificeret ved hjælp af ImageJ, og data er gennemsnit ± SEM for 10-12 mus pr. gruppe (b). * p <0,01 eller *** p <0,0001 vs kontrol (C) og # p <0,01 eller # # p <0,001 vs den respektive dosis metamfetamin (Tukey's multiple sammenligningstest).
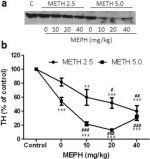
Fig. 3a viser, at mephedron signifikant øgede metamfetamininducerede reduktioner i TH-niveauer som bestemt ved immunoblotting. Immunoblots blev kvantificeret, og i overensstemmelse med resultaterne ovenfor for DA og DAT var hovedeffekterne af metamfetamindosis (F1,81 = 47.89, p <0.0001) og mephedronedosis (F4,81 = 63.57, p <0.0001) meget signifikante ved tovejs ANOVA (Fig. 3b). Hovedeffekten af mephedron givet i kombination med enten 2,5 mg / kg (F4,34 = 12,98, p < 0,0001) eller 5,0 mg / kg metamfetamin (F4,49 = 99,16, p < 0,0001) var også meget signifikant ved envejs ANOVA. Alle behandlinger med begge doser af metamfetamin ± mefedron forårsagede signifikant større reduktioner i TH sammenlignet med den respektive kontrol (p < 0,001 for 2,5 mg/kg metamfetamin + 10 mg/kg mefedron; p < 0,0001 for alle andre kombinationer) med undtagelse af 2,5 mg/kg metamfetamin alene, som ikke ændrede TH-niveauerne signifikant (dvs. ingen toksicitet). Fig. 3b viser også, at mephedron-doser på 20 mg/kg (p < 0,01) og 40 mg/kg (p < 0,001) signifikant forstærkede reduktionerne i TH forårsaget af 2,5 mg/kg metamfetamin, og at alle tre doser mephedron signifikant (p < 0,0001) forstærkede virkningerne af 5,0 mg/kg metamfetamin på TH-reduktionerne.
Fig. 3
Effekter af mephedron på metamfetamininducerede reduktioner i striatal TH. Mus blev behandlet med de angivne doser af mephedron (MEPH) 30 minutter før hver injektion af 2,5 (●) eller 5,0 mg / kg (■) metamfetamin (METH) og ofret 2d senere til bestemmelse af striatal niveauer af TH ved immunoblotting (a). Blots blev kvantificeret ved hjælp af ImageJ, og data er gennemsnit ± SEM for 10-12 mus pr. gruppe (b). Nogle fejlbjælker var for små til at overskride størrelsen på symbolerne og vises ikke synlige. **p < 0,001 eller ***p < 0,0001 vs kontrol (C) og #p < 0,01, ##p < 0,001 eller ###p < 0,0001) vs den respektive dosis metamfetamin (Tukey's multiple sammenligningstest).
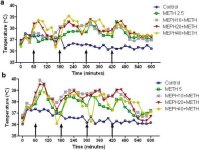
Virkninger af mephedron på metamfetamininduceret hypertermi
Mephedron forårsager ligesom metamfetamin betydelig hypertermi (Hadlock et al. 2011, Baumann et al. 2012, Angoa-Perez et al. 2012). Når mefedron blev givet 30 minutter før hver injektion af metamfetamin, kan det ses i fig. 4, at hovedeffekterne af metamfetamin- og mefedrondoser (F1,300 = 11,99, p < 0,0001) over tid (F4,300 = 51,73, p < 0,0001) var meget signifikante ved tovejs ANOVA. Hovedeffekterne af mephedron givet i kombination med enten 2,5 mg/kg metamfetamin (F4,120 = 41,44, p < 0,0001, panel a) over tid (F30,120 = 3,84, p < 0,0001) eller 5,0 mg/kg metamfetamin (F4,120 = 78,09, p < 0,0001, panel b) over tid (F30,120 = 9,98, p < 0,0001) var også meget signifikante ved tovejs ANOVA. Alle behandlinger med begge doser af metamfetamin ± mephedron var signifikant forskellige fra de respektive kontroller (p < 0,0001 for alle behandlinger).
Fig. 4
Effekter af mephedron på metamfetamin-induceret hypertermi. Mus blev behandlet med de angivne doser af mephedron (MEPH) 30 minutter før hver injektion af 2,5 (a) eller 5,0 mg/kg (b) metamfetamin (METH). Kernetemperaturer blev målt med 20 minutters mellemrum ved hjælp af telemetri, der startede 60 minutter før den første injektion af metamfetamin. De 4 metamfetamininjektioner er angivet med pilene, der hviler på x-aksen. Data udtrykkes som gennemsnitlig kropstemperatur for 6-8 mus pr. gruppe. SEM'er var altid < 10% af gennemsnittet og er udeladt for klarhedens skyld.
Virkninger af mephedron på amfetamin- og MDMA-induceret neurotoksicitet
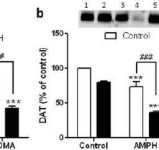
For at teste, om de forstærkende virkninger af mephedron på metamfetamin kunne udvides til andre neurotoksiske amfetaminer, blev mus behandlet med denne β-ketoamfetamin (20 mg/kg) plus amfetamin (4X 5 mg/kg) eller MDMA (4X 20 mg/kg), og resultaterne er præsenteret i fig. 5. Husk, at mephedron i sig selv ikke reducerer striatal DA, DAT eller TH (Angoa-Perez et al. 2012). Hovedvirkningen af lægemiddel (F5,27 = 27.18, p <0.0001) var meget signifikant ved envejs ANOVA for DA-reduktioner (fig. 5a). Det kan også ses i fig. 5a, at alle behandlinger med amfetamin (p <0.001) eller MDMA (p <0.001) alene eller i kombination med mephedron (p <0.0001 for begge lægemidler) signifikant sænkede DA-niveauer fra kontrol. Mephedron forbedrede signifikant DA-reduktioner forårsaget af amfetamin (p <0.01) eller MDMA (p <0.01). Fig. 5b viser lignende effekter af kombinationslægemiddelbehandlinger på DAT-niveauer i striatum. Hovedvirkningen af medikament (F4,49 = 42.63, p <0.0001) var meget signifikant ved envejs ANOVA for DAT. Det kan også ses i fig. 5b, at alle behandlinger med amfetamin eller MDMA var signifikant (p <0.0001 for alle) lavere sammenlignet med kontrol. Mephedrone forbedrede også signifikant DAT-reduktioner forårsaget af enten amfetamin eller MDMA (p <0.0001 i begge tilfælde). Endelig Fig. 5c viser, at lægemidlets hovedeffekt (F4,50 = 75.06, p <0.0001) var meget signifikant ved envejs ANOVA for reduktioner i TH. Det kan også ses i fig. 5c, at alle behandlinger med amfetamin eller MDMA var signifikant (p < 0,0001 for alle) lavere sammenlignet med kontrol. Mephedrone forbedrede også signifikant TH-reduktioner forårsaget af enten amfetamin eller MDMA (p <0.0001 i begge tilfælde)
Fig. 5
Effekter af mephedron på amfetamin- eller MDMA-induceret DA-nerveafsluttende neurotoksicitet. Mus blev behandlet med 20 mg / kg mephedron (MEPH) 30 minutter før hver injektion af 5,0 mg / kg amfetamin (AMPH) eller 20 mg / kg MDMA og ofret 2d efter behandling til bestemmelse af striatal niveauer af (a) DA ved HPLC. (b) DAT og (c) TH blev bestemt ved immunoblotting, og blots blev kvantificeret ved hjælp af ImageJ. Repræsentative immunoblots for DAT og TH er inkluderet som indsatser i henholdsvis paneler (b) og (c), og behandlinger for begge paneler er angivet med 1,5: kontrol; 2,6: MEPH; 3: AMPH; 4: AMPH + MEPH; 7: MDMA; og 8: MDMA + MEPH. Data er gennemsnit ± SEM for 5-12 mus i hver gruppe. **p <0,001 eller ***p <0,0001 vs kontrol og #p <0,01 eller ###p <0,0001 vs AMPH eller MDMA (Tukey's multiple sammenligningstest).
Effekter af nomifensin på metamfetamininduceret neurotoksicitet
Nomifensine, en kraftig DAT-blokker uden kendt misbrug eller neurotoksisk potentiale, blev testet for sin evne til at beskytte mod metamfetamininduceret neurotoksicitet og for kontrast til mephedrons handlinger på toksiciteten for DA-nerveender forårsaget af metamfetamin, amfetamin og MDMA. Resultaterne i fig. 6a viser, at hovedvirkningen af lægemiddel (F3,16 = 63,39, p < 0,0001) på DA-niveauer var meget signifikant ved envejs ANOVA. Nomifensin alene ændrede ikke DA-niveauer, men reduktionen forårsaget af metamfetamin (p < 0,0001) blev let, men signifikant vendt af nomifensin (p < 0,01). Hovedvirkningen af medikament (F3,20 = 16.78, p <0.0001) på DAT-niveauer var meget signifikant ved envejs ANOVA som vist i Fig. 6b. Nomifensin ændrede ikke DAT-niveauer, men gav signifikant beskyttelse (p <0.001) mod reduktionen i striatal DAT forårsaget af methamfetamin (p <0.0001) i sammenligning med kontrol. Endelig Fig. 6c viser, at hovedvirkningen af medikament (F3,15 = 14.10, p <0.0001) på TH-niveauer var meget signifikant ved envejs ANOVA. Som det ses for DA og DAT, blev reduktionen i TH forårsaget af metamfetamin (p <0.0001) let, men signifikant forhindret af nomifensin (p <0.01).
Fig. 6
Effekter af nomifensin på metamfetamin-induceret DA-nerveende neurotoksicitet. Mus blev behandlet med 5,0 mg / kg nomifensin (NOM) 30 minutter før hver injektion af 5,0 mg / kg methamphetamin (METH) og ofret 2d senere til bestemmelse af striatal niveauer af (a) DA ved HPLC. (b) DAT og (c) TH blev bestemt ved immunoblotting, og blots blev kvantificeret ved hjælp af ImageJ. Repræsentative immunoblots for DAT og TH er inkluderet som indsatser i panelerne (b) og (c) respektfuldt. Data er gennemsnit plus SEM for 5-7 mus pr. Gruppe. ***p <0.0001 vs kontrol (C) og #p <0.01 eller ##p <0.001 vs metamfetamin alene (Tukey's multiple sammenligningstest).
Diskussion
Formålet med denne undersøgelse var at afgøre, om mephedron ville forhindre DA-nerveende toksicitet forårsaget af methamphetamin. Baseret på dets kemiske lighed med metamfetamin og MDMA var det oprindeligt forventet, at mephedron ville udøve skadelige virkninger på DA-neuronsystemet. Men flere undersøgelser fastslog næsten samtidig, at mephedron ikke var giftigt for DA-nerveender (Angoa-Perez et al. 2012, Baumann et al. 2012, Hadlock et al. 2011). Spørgsmålet om, hvorvidt dette stof forårsager skade på det neuronale 5-HT-system, står stadig åbent. En undersøgelse rapporterede vedvarende reduktioner i 5-HT-nerveendernes funktion (Hadlock et al. 2011), mens en anden fandt, at mephedron ikke forårsagede skade (Baumann et al. 2012). Mephedron interagerer med DA-nerveafslutningen på en måde, der tyder på, at det faktisk stimulerer frigivelse og blokerer DA-genoptagelse via dets interaktioner med DAT. En vigtig del af metamfetamins neurotoksiske virkningsmekanisme er dets evne til at få adgang til DA-nerveender gennem DAT og forstyrre DA-homøostasen (Sulzer 2011). Hvis dette tidlige trin i metamfetamins neurotoksiske kaskade forhindres ved at hæmme DAT, forhindres toksicitet (Pu et al. 1994, Poth et al. 2012, Marek et al. 1990, Schmidt og Gibb 1985). Vi tænkte, at mephedron kunne have den samme beskyttende egenskab som andre DAT-hæmmere, men observerede i stedet en betydelig forøgelse af toksiciteten. Denne interaktion blev set ved hjælp af to forskellige doser metamfetamin, der forårsager moderat eller alvorlig skade på DA-nerveender (henholdsvis 4X 2,5 eller 5,0 mg/kg). Denne forstærkende effekt af mephedron var ikke begrænset til metamfetamin, men omfattede også amfetamin og MDMA, to stoffer, der ofte misbruges sammen med mephedron og andre β-ketoamfetaminer (Feyissa og Kelly 2008, Schifano et al. 2011, Kelly 2011). På trods af, at mephedron ikke forårsager toksicitet for i det mindste DA-nerveender i striatum, forstærker det derfor de neurotoksiske virkninger af andre misbrugsstoffer. Dette nye fund bør sætte mephedronmisbrug i et endnu mere grelt lys, fordi dets manglende iboende neurotoksicitet kan få det til at virke uskadeligt.
Hypertermi er en almindeligt rapporteret akut bivirkning ved indtagelse af metamfetamin (Greene et al. 2008) og β-ketoamfetamin hos mennesker (Borek og Holstege 2012, Prosser og Nelson 2012). Ligesom metamfetamin forårsager mange af β-ketoamfetamin-stofferne også betydelige stigninger i kernetemperaturen hos gnavere (Angoa-Perez et al. 2012, Hadlock et al. 2011, Baumann et al. 2012, Rockhold et al. 1997). Mens hypertermi forårsaget af metamfetamin kan bidrage til dets morfologiske og neuronale skadevirkninger, er det ikke nødvendigvis tilfældet, at hypertermi er den direkte årsag til disse virkninger (Kiyatkin og Sharma 2009). Vi registrerede kropstemperaturen hos mus, der blev behandlet med mephedron og metamfetamin, og observerede, at den kombinerede behandling ikke øgede temperaturen ud over de maksimale stigninger, der blev set efter begge stoffer alene. Metamfetamin forårsagede en dosisrelateret stigning i kropstemperaturen, og denne hypertermi var uforanderlig over hele det testede mephedron-dosisområde. Faktisk blev faldet i kropstemperatur efter injektion observeret efter mephedronbehandling (Angoa-Perez et al. 2012) bibeholdt ved højere doser af mephedron plus metamfetamin. Selv om den lægemiddelinducerede hypertermi ikke blev forstærket af kombineret lægemiddelbehandling, var de neurotoksiske virkninger additive. Derfor ser det i det mindste i dette tilfælde ud til, at de neurotoksiske virkninger af metamfetamin kan forstærkes af mephedron på en måde, der er uafhængig af hypertermi.
Mephedron hæmmer klart DAT-funktionen og blokerer DA-genoptagelse in vitro (Lopez-Arnau et al. 2012, Hadlock et al. 2011, Kehr et al. 2011, Martinez-Clemente et al. 2012, Cozzi et al. 1999). Mephedron fortrænger WIN-35,428 fra dets bindingssted på DAT, hvilket tyder på, at det er en konkurrencedygtig hæmmer af DA-optagelsen (Martinez-Clemente et al. 2012, Lopez-Arnau et al. 2012). Mephedrons styrke i denne henseende er meget lig metamfetamins (Cozzi et al. 1999) og MDMA's (Escubedo et al. 2011). Det vides ikke, om mephedron transporteres af DAT, men det gør methcathinon (Cozzi og Foley 2003). Nomifensin og amfonsyre, som binder sig til DAT og hæmmer DA-optagelsen, giver betydelig beskyttelse mod metamfetamin-induceret neurotoksicitet (Pu et al. 1994, Marek et al. 1990, Schmidt og Gibb 1985, Poth et al. 2012), og mus, der mangler DAT, er resistente over for metamfetamins neuronale toksicitet (Fumagalli et al. 1998). Når man ved, at mephedron ikke er neurotoksisk og er en DAT-blokker, kan man forudsige, at det bør forhindre toksicitet. Vi testede nomifensin i denne henseende som en positiv kontrol og bekræftede, at det beskytter mod metamfetamininduceret udtømning af DA, DAT og TH. Nomifensin hæmmer også noradrenalintransportøren (Brogden et al. 1979), men denne egenskab kan ikke forklare de nuværende resultater, fordi de fleste β-ketoamfetaminer, herunder mephedron, hæmmer noradrenalintransportøren og blokerer optagelsen af noradrenalin (Kelly 2011, Rothman et al. 2003, Cozzi et al. 1999, Sogawa et al. 2011, Lopez-Arnau et al. 2012). En rolle for det neurale 5-HT-system i nogle af de farmakologiske virkninger af mephedron er mulig i lyset af dette stofs evne til, ligesom MDMA (Yamamoto et al. 1995), at forårsage udstrømning af striatal DA via dets interaktioner med 5-HT2A-receptorer (Lopez-Arnau et al. 2012, Martinez-Clemente et al. 2012). Hyperlokomotion forårsaget af mephedron er afhængig af endogent 5-HT (Lopez-Arnau et al. 2012), og dette stof stimulerer også frigivelsen af 5-HT og hæmmer dets optagelse in vitro (Sogawa et al. 2011, Cozzi et al. 1999, Nagai et al. 2007, Hadlock et al. 2011, Lopez-Arnau et al. 2012, Martinez-Clemente et al. 2012) og in vivo (Baumann et al. 2012, Kehr et al. 2011). Vi kan dog udelukke en rolle for endogent 5-HT i DA-neurotoksiciteten af i det mindste metamfetamin ved at vise, at mus, der er genetisk udtømt for 5-HT, bevarer deres følsomhed over for neurotoksicitet (Thomas et al. 2010).
Mephedron kan øge metamfetamins neurotoksicitet ved hjælp af flere mulige mekanismer. For det første kan mephedron interagere med VMAT for at forårsage lækage af DA i cytoplasmaet i den præsynaptiske nerveende. Behandlinger, der øger den cytoplasmatiske pulje (dvs. lægemiddelfrigørelse) af DA, øger metamfetamin-neurotoksiciteten (Thomas et al. 2008, Thomas et al. 2009, Schmidt et al. 1985). Denne mekanisme er ikke sandsynlig, fordi methcathinon kun interagerer svagt med VMAT (Cozzi et al. 1999). For det andet kunne kombinationen af mephedron plus metamfetamin have en synergistisk effekt på ikke-vesikulær frigivelse af DA, men denne mulighed virker også usandsynlig i lyset af resultater, der viser, at behandling af DAT- eller SERT-udtrykkende CHO-celler med methylon plus metamfetamin ikke har en additiv effekt på frigivelse af DA eller 5-HT (Sogawa et al. 2011). For det tredje kan mephedron interagere med DAT på en ny måde, der bidrager til additiv toksicitet. Det er blevet påvist, at methylon i kombination med metamfetamin forårsager synergistisk cytotoksicitet i CHO-celler, der udtrykker DAT eller SERT, men ikke i vildtype CHO-celler, der mangler transportørerne (Sogawa et al. 2011). Den cytotoksicitet, der ses i dyrkede celler i disse undersøgelser (dvs. LDH-frigivelse), er meget forskellig fra den skade på DA-nerveender, der forårsages af metamfetamin, men denne mekanisme antyder en interessant, men udefineret rolle for DAT i øget cytotoksicitet. Endelig kan mephedron ændre metabolismen af metamfetamin. Mephedron metaboliseres primært ved N-demethylering (Meyer og Maurer 2010), ligesom metamfetamin og MDMA (Caldwell 1976). Denne mekanisme understøttes af påvisningen af, at metamfetamin og MDMA gensidigt hæmmer produktionen af deres respektive primære metabolitter og hæver stoffets plasmaniveauer over dem, der ses efter indgift af et af stofferne alene (Kuwayama et al. 2012). De doser af mephedron, der er brugt nu og i vores tidligere undersøgelse (Angoa-Perez et al. 2012), er ganske vist høje, men ikke neurotoksiske og ligger inden for det område, der misbruges af mennesker (McErath og O'Neill 2011). Derfor kan mephedron virke som MDMA og øge plasmaniveauet af metamfetamin ved at hæmme dets metabolisme. En dybdegående farmakokinetisk analyse vil være nødvendig for at bekræfte denne sidste mulighed.
Misbrug af β-ketoamfetaminer stiger med alarmerende hast, og mephedron er nu et af de mest anvendte stoffer efter cannabis, MDMA og kokain (Morris 2010, Winstock et al. 2011b). Desuden fremkalder mephedron stærkere følelser af trang hos mennesker sammenlignet med MDMA (Brunt et al. 2011), og brugere, der sniffer mephedron, vurderer det som mere vanedannende end kokain (Winstock et al. 2011b). Mephedron indtages af mennesker på en binge-lignende måde (dvs. "stacking") og tages ofte sammen med andre stoffer som cannabis og amfetamin-psykostimulanter (Schifano et al. 2011, Fass et al. 2012, Winstock et al. 2011a, Kelly 2011, Torrance and Cooper 2010). Mephedron findes i stigende grad i tabletter, der sælges som MDMA (Brunt et al. 2011), og brugen af det vil sandsynligvis overgå brugen af MDMA, da renheden af sidstnævnte stof fortsat falder (Brunt et al. 2011, Tanner-Smith 2006, Teng et al. 2006). På baggrund af de almindelige mønstre for misbrug af mephedron og andre "badesalt"-ingredienser er det vigtigt at overveje, om der opstår yderligere sundhedsrisici hos mennesker, når disse stoffer kombineres med amfetamin, bevidst eller ubevidst. Vores resultater, der viser, at mephedron i det mindste øger neurotoksiciteten for DA-nerveender i striatum forårsaget af metamfetamin, amfetamin og MDMA, afslører en særlig farlig og uventet egenskab ved denne β-ketoamfetamin.
Anvendte forkortelser
5-HT serotonin
DA dopamin
DAT DA-transportør
MDMA 3,4-methylendioxymetamfetamin
TH tyrosin hydroxylase
VMAT vesikulær monoamintransportør
Mephedron (4-methylmethcathinon) er et cathinonderivat og en strukturel analog til metamfetamin og 3,4-methylendioxy-metamfetamin (MDMA). Mefedron er en psykoaktiv ingrediens i "badesalte" sammen med andre forbindelser som methylon, butylon og 3,4-methylendioxypyrovaleron (MDPV). β-ketoamfetaminer misbruges i stigende grad, hvilket ikke mindst skyldes den meget begrænsede tilgængelighed af prækursorer, der er nødvendige for at syntetisere metamfetamin og MDMA i hemmelige laboratorier, og en tilsvarende reduktion af deres renhed (Winstock et al. 2011b, Brunt et al. 2011). I takt med at misbruget af β-ketoamfetamin fortsætter med at stige, er listen over skadevirkninger vokset til at omfatte kardiovaskulære komplikationer, uro, søvnløshed, psykose og depression (Schifano et al. 2011, Prosser og Nelson 2012).
Som kemisk beslægtet med metamfetamin og MDMA er det ikke overraskende, at β-ketoamfetaminerne har mange af de samme virkninger på centralnervesystemet som de førstnævnte stoffer. For eksempel blokerer disse stoffer dopamin- (DA) og serotonin- (5-HT) transportører (henholdsvis DAT og SERT) (Cozzi et al. 1999, Rothman et al. 2003, Fleckenstein et al. 2000, Lopez-Arnau et al. 2012), og de stimulerer frigivelse af monoaminer in vitro (Kalix og Glennon 1986, Gygi et al. 1997, Rothman et al. 2003) og in vivo (Gygi et al. 1997, Kehr et al. 2011). Methcathinone forårsager vedvarende reduktioner af tryptophan hydroxylase og tyrosin hydroxylase (TH) aktivitet og udtømning af DA og 5-HT (Gygi et al. 1997, Gygi et al. 1996, Sparago et al. 1996). PET-billeddannelsesundersøgelser hos afholdende methcathinonbrugere afslørede reduceret striatal DAT-tæthed, hvilket tyder på et tab af DA-terminaler (McCann et al. 1998). Den samtidige stimulering af DA-frigivelse og hæmning af optagelsen afspejler de kritiske elementer, der ligger til grund for den neurotoksicitet, der er forbundet med metamfetamin (Kuhn et al. 2008, Yamamoto og Bankson 2005, Cadet et al. 2007, Fleckenstein et al. 2007).
Vi (Angoa-Perez et al. 2012) og andre (Baumann et al. 2012, Hadlock et al. 2011) undersøgte for nylig muligheden for, at mephedron kunne forårsage neurotoksicitet ligesom metamfetamin og MDMA. Overraskende nok var mephedron ikke giftigt for DA-nerveender i striatum (Hadlock et al. 2011, Baumann et al. 2012, Angoa-Perez et al. 2012). Spørgsmålet om, hvorvidt mephedron skader 5-HT-nerveender, er stadig uafklaret, da en undersøgelse dokumenterede positive effekter (Hadlock et al. 2011), mens en anden var negativ (Baumann et al. 2012). I lyset af den relativt godartede effekt af mephedron på DA-nerveender og i betragtning af dets egenskaber som DAT-blokker, antog vi, at det faktisk kunne beskytte DA-neuronsystemet mod de neurotoksiske virkninger af metamfetamin, ligesom det vides at forekomme med andre DAT-blokkere såsom amfonsyre (Pu et al. 1994, Schmidt og Gibb 1985, Marek et al. 1990) og nomifensin (Poth et al. 2012). Vi rapporterer nu, at mephedron øger metamfetamins neurotoksicitet betydeligt. Denne effekt gælder også for amfetamin og MDMA, stoffer, der ofte misbruges sammen med mephedron (Feyissa og Kelly 2008, Schifano et al. 2011). Disse overraskende resultater sætter misbruget af mephedron i et nyt lys og gør det endnu mere presserende at få øjnene op for denne subtile og farlige egenskab ved β-ketoamfetamin.
Materialer og metoder
Stoffer og reagenser
Mephedronhydrochlorid og 3,4-methylendioxymetamfetamin (MDMA) blev anskaffet fra NIDA Research Resources Drug Supply Program. (+) Metamfetaminhydrochlorid, nomifensinmaleat, d-amfetaminsulfat, pentobarbital, DA og alle buffere og HPLC-reagenser blev købt fra Sigma-Aldrich (St. Louis, MO, USA). Bicinchoninic acid protein assay kits blev købt hos Pierce (Rockford, IL, USA). Polyklonale antistoffer mod TH fra rotter blev fremstillet som tidligere beskrevet (Kuhn og Billingsley 1987). Monoklonale antistoffer mod rotte-DAT blev generøst leveret af Dr. Roxanne A. Vaughan (University of North Dakota, Grand Forks, ND, USA). HRP-konjugerede anti-IgG sekundære antistoffer blev leveret af Jackson ImmunoResearch Laboratories, Inc. (West Grove, PA, USA).
Dyrene
C57BL/6-hunnemus (Harlan, Indianapolis, IN, USA), der vejede 20-25 g på forsøgstidspunktet, blev anbragt 5 pr. bur i store skotøjsbure i et lyst (12 timer lys/mørk) og temperaturkontrolleret rum. Der blev brugt hunmus, fordi de er kendt for at være meget følsomme over for neuronale skader af de neurotoksiske amfetaminer og for at opretholde konsistensen med vores tidligere undersøgelser af metamfetamins neurotoksicitet (Thomas et al. 2010, Thomas et al. 2008, Thomas et al. 2009). Musene havde fri adgang til mad og vand. Udvalget for institutionel pleje og brug af Wayne State University godkendte dyreplejen og de eksperimentelle procedurer. Alle procedurer var også i overensstemmelse med NIH Guide for the Care and Use of Laboratory Animals.
Farmakologiske, fysiologiske og adfærdsmæssige procedurer
Mus blev behandlet med mephedron ved hjælp af et binge-lignende regime bestående af 4 injektioner på 10, 20 eller 40 mg / kg med et 2 timers interval mellem hver injektion. Denne binge-behandling, når den bruges til at injicere substituerede amfetaminer og katinonderivater, resulterer i omfattende DA-nerveskader. De doser af mephedron, der anvendes i øjeblikket, har tidligere vist sig ikke at være giftige for DA-nerveender (Angoa-Perez et al. 2012). Mus blev behandlet med metamfetamin (4X 2,5 eller 5 mg/kg), amfetamin (4X 5 mg/kg) eller MDMA (4X 20 mg/kg) alene eller i kombination med mephedron. Når musene blev behandlet med to stoffer, fik de en mephedron-injektion 30 minutter før hver af de fire injektioner af metamfetamin, amfetamin eller MDMA. Kontrolgruppen fik indsprøjtninger med fysiologisk saltvand på samme tidspunkt som for mephedron alene eller i kombination med andre amfetaminer. Som kontrol for virkningerne af en DAT-hæmmer på metamfetamintoksicitet blev musene behandlet med nomifensin (4X 5 mg/kg) 30 minutter før hver injektion af metamfetamin (4X 5 mg/kg). Alle indsprøjtninger blev givet via i.p.-ruten. Musene blev ofret 2 dage efter den sidste lægemiddelbehandling, når den amfetamin-associerede neurotoksicitet havde nået sit maksimum. Kropstemperaturen blev overvåget ved hjælp af telemetri med IPTT-300 implanterbare temperaturtranspondere fra Bio Medic Data Systems, Inc. (Seaford, DE, USA). Temperaturerne blev registreret ikke-invasivt hvert 20. minut, startende 60 minutter før den første METH-injektion og fortsatte i 9 timer derefter ved hjælp af DAS-5001-konsolsystemet fra Bio Medic.
Bestemmelse af striatal DA-indhold
Striatalvæv blev dissekeret bilateralt fra hjernen efter behandling og opbevaret ved -80 ° C. Frosset væv blev vejet og sonikeret i 10 volumener 0,16 N perchlorsyre ved 4 °C. Uopløseligt protein blev fjernet ved centrifugering, og DA blev bestemt ved HPLC med elektrokemisk detektion som tidligere beskrevet for metamfetamin (Thomas et al. 2010, Thomas et al, 2009).
Bestemmelse af TH- og DAT-proteinniveauer ved immunoblotting
Virkningerne af lægemiddelbehandlinger på striatal TH- og DAT-niveauer blev bestemt ved immunoblotting som et indeks for toksicitet for striatal DA-nerveender. Mus blev ofret ved halshugning efter behandling, og striatum blev dissekeret bilateralt. Vævet blev opbevaret ved -80 °C. Frosset væv blev opløst ved sonikering i 1 % SDS ved 95 °C, og uopløseligt materiale blev sedimenteret ved centrifugering. Protein blev bestemt ved hjælp af bicinchoninsyremetoden, og lige store mængder protein (70 μg/lane) blev opløst ved SDS-polyacrylamidgelelektroforese og derefter elektroblottet til nitrocellulose. Blots blev blokeret i Tris buffered saline indeholdende Tween 20 (0,1% v/v) og 5% fedtfri tørmælk i 1 time ved stuetemperatur. Primære antistoffer mod TH (1:1000) eller DAT (1:1000) blev tilsat til blots og fik lov til at inkubere i 16 timer ved 4 °C. Blots blev vasket 3 gange i Tris-bufferet saltvand for at fjerne ureagerede antistoffer og blev derefter inkuberet med HRP-konjugeret anti-IgG sekundært antistof (1:4000) i 1 time ved stuetemperatur. Immunreaktive bånd blev visualiseret ved forbedret kemiluminescens, og de relative tætheder af TH- og DAT-reaktive bånd blev bestemt ved billeddannelse med en Kodak Image Station (Carestream Molecular Systems, Rochester, NY, USA) og kvantificeret ved hjælp af ImageJ-software (NIH).
Analyse af data
To-vejs ANOVA'er blev udført for at analysere dosisvirkningerne af methamphetamin versus mephedron på DA, DAT og TH. Virkningerne af lægemiddelbehandlinger på striatal DA, TH og DAT-indhold blev testet for signifikans ved envejs ANOVA efterfulgt af Tukey's multiple sammenligningstest. Resultater af lægemiddelbehandlinger på kernekropstemperatur over tid blev analyseret ved hjælp af en tovejs ANOVA efterfulgt af Bonferronis test for at bestemme betydningen af forskelle i temperatur på individuelle tidspunkter efter behandling. Forskelle blev betragtet som signifikante, hvis p < 0,05. Alle statistiske analyser blev udført ved hjælp af GraphPad Prism version 5.02 til Windows (GraphPad Software, San Diego, CA, USA, www.graphpad.com).
Gå til:
Resultater
Virkninger af mephedron på metamfetamin-induceret neurotoksicitet
Mephedron i doser (10, 20 eller 40 mg/kg), der vides ikke at forårsage toksicitet i DA-nerven (Angoa-Perez et al. 2012), blev administreret 30 minutter før hver injektion af metamfetamin. Metamfetamin blev givet i doser, der forårsager moderat (4X 2,5 mg/kg) eller alvorlig (4X 5 mg/kg) skade på DA-nerveender i striatum (Thomas et al. 2004, Thomas et al. 2010). Resultater præsenteret i fig. 1 viser, at hovedeffekterne af metamfetamindosis (F1,40 = 66,60, p < 0,0001) og mephedron-dosis (F4,40 = 131,3, p < 0,0001) på DA-niveauer i striatum var meget signifikante ved tovejs ANOVA. Hovedeffekten af mephedron givet i kombination med enten 2.5 mg / kg (F4,22 = 35.96, p <0.001) eller 5.0 mg / kg methamphetamin (F4,17 = 953.9, p <0.0001) var også meget signifikant ved envejs ANOVA. Alle behandlinger med enten dosis metamfetamin ± mephedron forårsagede signifikant større reduktioner i DA sammenlignet med den respektive kontrol (p <0.0001 for alle). Fig. 1 viser også, at mephedron-doser på 20 (p < 0,01) og 40 mg/kg (p < 0,001) signifikant forstærkede de nedbrydende virkninger af 2,5 mg/kg metamfetamin på DA, mens alle doser af mephedron signifikant forstærkede virkningerne af 5,0 mg/kg metamfetamin på DA-niveauer (p < 0,0001 for alle).
http://bbzzzsvqcrqtki6umym6itiixfhni37ybtt7mkbjyxn2pgllzxf2qgyd.onion/index.php?attachments/sywobkqrnd-jpg.47/&hash=f31155ada4e59f22a35f9110aaa6a7e5
Fig.1
Virkninger af mephedron på metamfetamininducerede reduktioner i striatal DA. Mus blev behandlet med de angivne doser af mephedron (MEPH) 30 minutter før hver injektion af 2.5 (-) eller 5.0 mg / kg (■) methamphetamin (METH) og ofret 2d senere til bestemmelse af striatal niveauer af DA ved HPLC. Data er gennemsnit ± SEM for 5-7 mus pr. Gruppe. Nogle fejlbjælker var for små til at overstige størrelsen på symbolerne og vises ikke synlige. ***p < 0,001 vs kontroller og #p < 0,01, ##p < 0,001 eller ###p < 0,0001 vs den respektive dosis metamfetamin (Tukey's multiple sammenligningstest).
Fig. 2a viser, at mephedron signifikant øgede metamfetamininducerede reduktioner i DAT-niveauer som bestemt ved immunoblotting. Immunoblots blev kvantificeret, og i overensstemmelse med resultaterne for DA var hovedeffekterne af metamfetamindosis (F1,92 = 9.48, p <0.001) og mephedronedosis (F4,92 = 37.56, p <0.0001) på DAT-niveauer i striatum meget signifikante ved tovejs ANOVA (Fig. 2b). Hovedeffekten af mephedron givet i kombination med enten 2.5 mg / kg (F4,56 = 15.55, p <0.0001) eller 5.0 mg / kg metamfetamin (F4,39 = 24.84, p <0.0001) var også meget signifikant ved envejs ANOVA. Alle behandlinger med enten dosis metamfetamin ± mephedron forårsagede signifikant større reduktioner i DAT sammenlignet med den respektive kontrol (p <0.01 for 2.5 mg / kg metamfetamin alene; p <0.0001 for alle andre behandlinger). Fig. 2b viser også, at mephedron-doser på 20 mg / kg (p <0.01) og 40 mg / kg (p <0.001) signifikant forbedrede reduktionerne i DAT forårsaget af 2.5 mg / kg methamphetamin, mens kun 40 mg / kg mephedron-dosis signifikant forbedrede (p <0.01) virkningerne af 5.0 mg / kg methamphetamin på DAT-reduktioner.
http://bbzzzsvqcrqtki6umym6itiixfhni37ybtt7mkbjyxn2pgllzxf2qgyd.onion/index.php?attachments/bfu3sx5dnr-jpg.4797/&hash=f31155ada4e59f22a35f9110aaa6a7e5
Fig.2
Effekter af mephedron på metamfetamin-inducerede reduktioner i striatal DAT. Mus blev behandlet med de angivne doser af mephedron (MEPH) 30 minutter før hver injektion af 2.5 (●) eller 5.0 mg / kg (■) metamfetamin (METH) og ofret 2d senere til bestemmelse af striatal niveauer af DAT ved immunoblotting (a). Blots blev kvantificeret ved hjælp af ImageJ, og data er gennemsnit ± SEM for 10-12 mus pr. gruppe (b). * p <0,01 eller *** p <0,0001 vs kontrol (C) og # p <0,01 eller # # p <0,001 vs den respektive dosis metamfetamin (Tukey's multiple sammenligningstest).
Fig. 3a viser, at mephedron signifikant øgede metamfetamininducerede reduktioner i TH-niveauer som bestemt ved immunoblotting. Immunoblots blev kvantificeret, og i overensstemmelse med resultaterne ovenfor for DA og DAT var hovedeffekterne af metamfetamindosis (F1,81 = 47.89, p <0.0001) og mephedronedosis (F4,81 = 63.57, p <0.0001) meget signifikante ved tovejs ANOVA (Fig. 3b). Hovedeffekten af mephedron givet i kombination med enten 2,5 mg / kg (F4,34 = 12,98, p < 0,0001) eller 5,0 mg / kg metamfetamin (F4,49 = 99,16, p < 0,0001) var også meget signifikant ved envejs ANOVA. Alle behandlinger med begge doser af metamfetamin ± mefedron forårsagede signifikant større reduktioner i TH sammenlignet med den respektive kontrol (p < 0,001 for 2,5 mg/kg metamfetamin + 10 mg/kg mefedron; p < 0,0001 for alle andre kombinationer) med undtagelse af 2,5 mg/kg metamfetamin alene, som ikke ændrede TH-niveauerne signifikant (dvs. ingen toksicitet). Fig. 3b viser også, at mephedron-doser på 20 mg/kg (p < 0,01) og 40 mg/kg (p < 0,001) signifikant forstærkede reduktionerne i TH forårsaget af 2,5 mg/kg metamfetamin, og at alle tre doser mephedron signifikant (p < 0,0001) forstærkede virkningerne af 5,0 mg/kg metamfetamin på TH-reduktionerne.
http://bbzzzsvqcrqtki6umym6itiixfhni37ybtt7mkbjyxn2pgllzxf2qgyd.onion/index.php?attachments/tggfrjpqru-jpg.4798/&hash=f31155ada4e59f22a35f9110aaa6a7e5
Fig. 3
Effekter af mephedron på metamfetamininducerede reduktioner i striatal TH. Mus blev behandlet med de angivne doser af mephedron (MEPH) 30 minutter før hver injektion af 2,5 (●) eller 5,0 mg / kg (■) metamfetamin (METH) og ofret 2d senere til bestemmelse af striatal niveauer af TH ved immunoblotting (a). Blots blev kvantificeret ved hjælp af ImageJ, og data er gennemsnit ± SEM for 10-12 mus pr. gruppe (b). Nogle fejlbjælker var for små til at overskride størrelsen på symbolerne og vises ikke synlige. **p < 0,001 eller ***p < 0,0001 vs kontrol (C) og #p < 0,01, ##p < 0,001 eller ###p < 0,0001) vs den respektive dosis metamfetamin (Tukey's multiple sammenligningstest).
Virkninger af mephedron på metamfetamininduceret hypertermi
Mephedron forårsager ligesom metamfetamin betydelig hypertermi (Hadlock et al. 2011, Baumann et al. 2012, Angoa-Perez et al. 2012). Når mefedron blev givet 30 minutter før hver injektion af metamfetamin, kan det ses i fig. 4, at hovedeffekterne af metamfetamin- og mefedrondoser (F1,300 = 11,99, p < 0,0001) over tid (F4,300 = 51,73, p < 0,0001) var meget signifikante ved tovejs ANOVA. Hovedeffekterne af mephedron givet i kombination med enten 2,5 mg/kg metamfetamin (F4,120 = 41,44, p < 0,0001, panel a) over tid (F30,120 = 3,84, p < 0,0001) eller 5,0 mg/kg metamfetamin (F4,120 = 78,09, p < 0,0001, panel b) over tid (F30,120 = 9,98, p < 0,0001) var også meget signifikante ved tovejs ANOVA. Alle behandlinger med begge doser af metamfetamin ± mephedron var signifikant forskellige fra de respektive kontroller (p < 0,0001 for alle behandlinger).
http://bbzzzsvqcrqtki6umym6itiixfhni37ybtt7mkbjyxn2pgllzxf2qgyd.onion/index.php?attachments/x0ik9hdwsw-jpg.4799/&hash=f31155ada4e59f22a35f9110aaa6a7e5
Fig. 4
Effekter af mephedron på metamfetamin-induceret hypertermi. Mus blev behandlet med de angivne doser af mephedron (MEPH) 30 minutter før hver injektion af 2,5 (a) eller 5,0 mg/kg (b) metamfetamin (METH). Kernetemperaturer blev målt med 20 minutters mellemrum ved hjælp af telemetri, der startede 60 minutter før den første injektion af metamfetamin. De 4 metamfetamininjektioner er angivet med pilene, der hviler på x-aksen. Data udtrykkes som gennemsnitlig kropstemperatur for 6-8 mus pr. gruppe. SEM'er var altid < 10% af gennemsnittet og er udeladt for klarhedens skyld.
Virkninger af mephedron på amfetamin- og MDMA-induceret neurotoksicitet
For at teste, om de forstærkende virkninger af mephedron på metamfetamin kunne udvides til andre neurotoksiske amfetaminer, blev mus behandlet med denne β-ketoamfetamin (20 mg/kg) plus amfetamin (4X 5 mg/kg) eller MDMA (4X 20 mg/kg), og resultaterne er præsenteret i fig. 5. Husk, at mephedron i sig selv ikke reducerer striatal DA, DAT eller TH (Angoa-Perez et al. 2012). Hovedvirkningen af lægemiddel (F5,27 = 27.18, p <0.0001) var meget signifikant ved envejs ANOVA for DA-reduktioner (fig. 5a). Det kan også ses i fig. 5a, at alle behandlinger med amfetamin (p <0.001) eller MDMA (p <0.001) alene eller i kombination med mephedron (p <0.0001 for begge lægemidler) signifikant sænkede DA-niveauer fra kontrol. Mephedron forbedrede signifikant DA-reduktioner forårsaget af amfetamin (p <0.01) eller MDMA (p <0.01). Fig. 5b viser lignende effekter af kombinationslægemiddelbehandlinger på DAT-niveauer i striatum. Hovedvirkningen af medikament (F4,49 = 42.63, p <0.0001) var meget signifikant ved envejs ANOVA for DAT. Det kan også ses i fig. 5b, at alle behandlinger med amfetamin eller MDMA var signifikant (p <0.0001 for alle) lavere sammenlignet med kontrol. Mephedrone forbedrede også signifikant DAT-reduktioner forårsaget af enten amfetamin eller MDMA (p <0.0001 i begge tilfælde). Endelig Fig. 5c viser, at lægemidlets hovedeffekt (F4,50 = 75.06, p <0.0001) var meget signifikant ved envejs ANOVA for reduktioner i TH. Det kan også ses i fig. 5c, at alle behandlinger med amfetamin eller MDMA var signifikant (p < 0,0001 for alle) lavere sammenlignet med kontrol. Mephedrone forbedrede også signifikant TH-reduktioner forårsaget af enten amfetamin eller MDMA (p <0.0001 i begge tilfælde)
http://bbzzzsvqcrqtki6umym6itiixfhni37ybtt7mkbjyxn2pgllzxf2qgyd.onion/index.php?attachments/onvypkgxvn-jpg.4800/&hash=f31155ada4e59f22a35f9110aaa6a7e5
Fig. 5
Effekter af mephedron på amfetamin- eller MDMA-induceret DA-nerveafsluttende neurotoksicitet. Mus blev behandlet med 20 mg / kg mephedron (MEPH) 30 minutter før hver injektion af 5,0 mg / kg amfetamin (AMPH) eller 20 mg / kg MDMA og ofret 2d efter behandling til bestemmelse af striatal niveauer af (a) DA ved HPLC. (b) DAT og (c) TH blev bestemt ved immunoblotting, og blots blev kvantificeret ved hjælp af ImageJ. Repræsentative immunoblots for DAT og TH er inkluderet som indsatser i henholdsvis paneler (b) og (c), og behandlinger for begge paneler er angivet med 1,5: kontrol; 2,6: MEPH; 3: AMPH; 4: AMPH + MEPH; 7: MDMA; og 8: MDMA + MEPH. Data er gennemsnit ± SEM for 5-12 mus i hver gruppe. **p <0,001 eller ***p <0,0001 vs kontrol og #p <0,01 eller ###p <0,0001 vs AMPH eller MDMA (Tukey's multiple sammenligningstest).
Effekter af nomifensin på metamfetamininduceret neurotoksicitet
Nomifensine, en kraftig DAT-blokker uden kendt misbrug eller neurotoksisk potentiale, blev testet for sin evne til at beskytte mod metamfetamininduceret neurotoksicitet og for kontrast til mephedrons handlinger på toksiciteten for DA-nerveender forårsaget af metamfetamin, amfetamin og MDMA. Resultaterne i fig. 6a viser, at hovedvirkningen af lægemiddel (F3,16 = 63,39, p < 0,0001) på DA-niveauer var meget signifikant ved envejs ANOVA. Nomifensin alene ændrede ikke DA-niveauer, men reduktionen forårsaget af metamfetamin (p < 0,0001) blev let, men signifikant vendt af nomifensin (p < 0,01). Hovedvirkningen af medikament (F3,20 = 16.78, p <0.0001) på DAT-niveauer var meget signifikant ved envejs ANOVA som vist i Fig. 6b. Nomifensin ændrede ikke DAT-niveauer, men gav signifikant beskyttelse (p <0.001) mod reduktionen i striatal DAT forårsaget af methamfetamin (p <0.0001) i sammenligning med kontrol. Endelig Fig. 6c viser, at hovedvirkningen af medikament (F3,15 = 14.10, p <0.0001) på TH-niveauer var meget signifikant ved envejs ANOVA. Som det ses for DA og DAT, blev reduktionen i TH forårsaget af metamfetamin (p <0.0001) let, men signifikant forhindret af nomifensin (p <0.01).
http://bbzzzsvqcrqtki6umym6itiixfhni37ybtt7mkbjyxn2pgllzxf2qgyd.onion/index.php?attachments/daczg9cwfv-jpg.4801/&hash=f31155ada4e59f22a35f9110aaa6a7e5
Fig. 6
Effekter af nomifensin på metamfetamin-induceret DA-nerveende neurotoksicitet. Mus blev behandlet med 5,0 mg / kg nomifensin (NOM) 30 minutter før hver injektion af 5,0 mg / kg methamphetamin (METH) og ofret 2d senere til bestemmelse af striatal niveauer af (a) DA ved HPLC. (b) DAT og (c) TH blev bestemt ved immunoblotting, og blots blev kvantificeret ved hjælp af ImageJ. Repræsentative immunoblots for DAT og TH er inkluderet som indsatser i panelerne (b) og (c) respektfuldt. Data er gennemsnit plus SEM for 5-7 mus pr. Gruppe. ***p <0.0001 vs kontrol (C) og #p <0.01 eller ##p <0.001 vs metamfetamin alene (Tukey's multiple sammenligningstest).
Diskussion
Formålet med denne undersøgelse var at afgøre, om mephedron ville forhindre DA-nerveende toksicitet forårsaget af methamphetamin. Baseret på dets kemiske lighed med metamfetamin og MDMA var det oprindeligt forventet, at mephedron ville udøve skadelige virkninger på DA-neuronsystemet. Men flere undersøgelser fastslog næsten samtidig, at mephedron ikke var giftigt for DA-nerveender (Angoa-Perez et al. 2012, Baumann et al. 2012, Hadlock et al. 2011). Spørgsmålet om, hvorvidt dette stof forårsager skade på det neuronale 5-HT-system, står stadig åbent. En undersøgelse rapporterede vedvarende reduktioner i 5-HT-nerveendernes funktion (Hadlock et al. 2011), mens en anden fandt, at mephedron ikke forårsagede skade (Baumann et al. 2012). Mephedron interagerer med DA-nerveafslutningen på en måde, der tyder på, at det faktisk stimulerer frigivelse og blokerer DA-genoptagelse via dets interaktioner med DAT. En vigtig del af metamfetamins neurotoksiske virkningsmekanisme er dets evne til at få adgang til DA-nerveender gennem DAT og forstyrre DA-homøostasen (Sulzer 2011). Hvis dette tidlige trin i metamfetamins neurotoksiske kaskade forhindres ved at hæmme DAT, forhindres toksicitet (Pu et al. 1994, Poth et al. 2012, Marek et al. 1990, Schmidt og Gibb 1985). Vi tænkte, at mephedron kunne have den samme beskyttende egenskab som andre DAT-hæmmere, men observerede i stedet en betydelig forøgelse af toksiciteten. Denne interaktion blev set ved hjælp af to forskellige doser metamfetamin, der forårsager moderat eller alvorlig skade på DA-nerveender (henholdsvis 4X 2,5 eller 5,0 mg/kg). Denne forstærkende effekt af mephedron var ikke begrænset til metamfetamin, men omfattede også amfetamin og MDMA, to stoffer, der ofte misbruges sammen med mephedron og andre β-ketoamfetaminer (Feyissa og Kelly 2008, Schifano et al. 2011, Kelly 2011). På trods af, at mephedron ikke forårsager toksicitet for i det mindste DA-nerveender i striatum, forstærker det derfor de neurotoksiske virkninger af andre misbrugsstoffer. Dette nye fund bør sætte mephedronmisbrug i et endnu mere grelt lys, fordi dets manglende iboende neurotoksicitet kan få det til at virke uskadeligt.
Hypertermi er en almindeligt rapporteret akut bivirkning ved indtagelse af metamfetamin (Greene et al. 2008) og β-ketoamfetamin hos mennesker (Borek og Holstege 2012, Prosser og Nelson 2012). Ligesom metamfetamin forårsager mange af β-ketoamfetamin-stofferne også betydelige stigninger i kernetemperaturen hos gnavere (Angoa-Perez et al. 2012, Hadlock et al. 2011, Baumann et al. 2012, Rockhold et al. 1997). Mens hypertermi forårsaget af metamfetamin kan bidrage til dets morfologiske og neuronale skadevirkninger, er det ikke nødvendigvis tilfældet, at hypertermi er den direkte årsag til disse virkninger (Kiyatkin og Sharma 2009). Vi registrerede kropstemperaturen hos mus, der blev behandlet med mephedron og metamfetamin, og observerede, at den kombinerede behandling ikke øgede temperaturen ud over de maksimale stigninger, der blev set efter begge stoffer alene. Metamfetamin forårsagede en dosisrelateret stigning i kropstemperaturen, og denne hypertermi var uforanderlig over hele det testede mephedron-dosisområde. Faktisk blev faldet i kropstemperatur efter injektion observeret efter mephedronbehandling (Angoa-Perez et al. 2012) bibeholdt ved højere doser af mephedron plus metamfetamin. Selv om den lægemiddelinducerede hypertermi ikke blev forstærket af kombineret lægemiddelbehandling, var de neurotoksiske virkninger additive. Derfor ser det i det mindste i dette tilfælde ud til, at de neurotoksiske virkninger af metamfetamin kan forstærkes af mephedron på en måde, der er uafhængig af hypertermi.
Mephedron hæmmer klart DAT-funktionen og blokerer DA-genoptagelse in vitro (Lopez-Arnau et al. 2012, Hadlock et al. 2011, Kehr et al. 2011, Martinez-Clemente et al. 2012, Cozzi et al. 1999). Mephedron fortrænger WIN-35,428 fra dets bindingssted på DAT, hvilket tyder på, at det er en konkurrencedygtig hæmmer af DA-optagelsen (Martinez-Clemente et al. 2012, Lopez-Arnau et al. 2012). Mephedrons styrke i denne henseende er meget lig metamfetamins (Cozzi et al. 1999) og MDMA's (Escubedo et al. 2011). Det vides ikke, om mephedron transporteres af DAT, men det gør methcathinon (Cozzi og Foley 2003). Nomifensin og amfonsyre, som binder sig til DAT og hæmmer DA-optagelsen, giver betydelig beskyttelse mod metamfetamin-induceret neurotoksicitet (Pu et al. 1994, Marek et al. 1990, Schmidt og Gibb 1985, Poth et al. 2012), og mus, der mangler DAT, er resistente over for metamfetamins neuronale toksicitet (Fumagalli et al. 1998). Når man ved, at mephedron ikke er neurotoksisk og er en DAT-blokker, kan man forudsige, at det bør forhindre toksicitet. Vi testede nomifensin i denne henseende som en positiv kontrol og bekræftede, at det beskytter mod metamfetamininduceret udtømning af DA, DAT og TH. Nomifensin hæmmer også noradrenalintransportøren (Brogden et al. 1979), men denne egenskab kan ikke forklare de nuværende resultater, fordi de fleste β-ketoamfetaminer, herunder mephedron, hæmmer noradrenalintransportøren og blokerer optagelsen af noradrenalin (Kelly 2011, Rothman et al. 2003, Cozzi et al. 1999, Sogawa et al. 2011, Lopez-Arnau et al. 2012). En rolle for det neurale 5-HT-system i nogle af de farmakologiske virkninger af mephedron er mulig i lyset af dette stofs evne til, ligesom MDMA (Yamamoto et al. 1995), at forårsage udstrømning af striatal DA via dets interaktioner med 5-HT2A-receptorer (Lopez-Arnau et al. 2012, Martinez-Clemente et al. 2012). Hyperlokomotion forårsaget af mephedron er afhængig af endogent 5-HT (Lopez-Arnau et al. 2012), og dette stof stimulerer også frigivelsen af 5-HT og hæmmer dets optagelse in vitro (Sogawa et al. 2011, Cozzi et al. 1999, Nagai et al. 2007, Hadlock et al. 2011, Lopez-Arnau et al. 2012, Martinez-Clemente et al. 2012) og in vivo (Baumann et al. 2012, Kehr et al. 2011). Vi kan dog udelukke en rolle for endogent 5-HT i DA-neurotoksiciteten af i det mindste metamfetamin ved at vise, at mus, der er genetisk udtømt for 5-HT, bevarer deres følsomhed over for neurotoksicitet (Thomas et al. 2010).
Mephedron kan øge metamfetamins neurotoksicitet ved hjælp af flere mulige mekanismer. For det første kan mephedron interagere med VMAT for at forårsage lækage af DA i cytoplasmaet i den præsynaptiske nerveende. Behandlinger, der øger den cytoplasmatiske pulje (dvs. lægemiddelfrigørelse) af DA, øger metamfetamin-neurotoksiciteten (Thomas et al. 2008, Thomas et al. 2009, Schmidt et al. 1985). Denne mekanisme er ikke sandsynlig, fordi methcathinon kun interagerer svagt med VMAT (Cozzi et al. 1999). For det andet kunne kombinationen af mephedron plus metamfetamin have en synergistisk effekt på ikke-vesikulær frigivelse af DA, men denne mulighed virker også usandsynlig i lyset af resultater, der viser, at behandling af DAT- eller SERT-udtrykkende CHO-celler med methylon plus metamfetamin ikke har en additiv effekt på frigivelse af DA eller 5-HT (Sogawa et al. 2011). For det tredje kan mephedron interagere med DAT på en ny måde, der bidrager til additiv toksicitet. Det er blevet påvist, at methylon i kombination med metamfetamin forårsager synergistisk cytotoksicitet i CHO-celler, der udtrykker DAT eller SERT, men ikke i vildtype CHO-celler, der mangler transportørerne (Sogawa et al. 2011). Den cytotoksicitet, der ses i dyrkede celler i disse undersøgelser (dvs. LDH-frigivelse), er meget forskellig fra den skade på DA-nerveender, der forårsages af metamfetamin, men denne mekanisme antyder en interessant, men udefineret rolle for DAT i øget cytotoksicitet. Endelig kan mephedron ændre metabolismen af metamfetamin. Mephedron metaboliseres primært ved N-demethylering (Meyer og Maurer 2010), ligesom metamfetamin og MDMA (Caldwell 1976). Denne mekanisme understøttes af påvisningen af, at metamfetamin og MDMA gensidigt hæmmer produktionen af deres respektive primære metabolitter og hæver stoffets plasmaniveauer over dem, der ses efter indgift af et af stofferne alene (Kuwayama et al. 2012). De doser af mephedron, der er brugt nu og i vores tidligere undersøgelse (Angoa-Perez et al. 2012), er ganske vist høje, men ikke neurotoksiske og ligger inden for det område, der misbruges af mennesker (McErath og O'Neill 2011). Derfor kan mephedron virke som MDMA og øge plasmaniveauet af metamfetamin ved at hæmme dets metabolisme. En dybdegående farmakokinetisk analyse vil være nødvendig for at bekræfte denne sidste mulighed.
Misbrug af β-ketoamfetaminer stiger med alarmerende hast, og mephedron er nu et af de mest anvendte stoffer efter cannabis, MDMA og kokain (Morris 2010, Winstock et al. 2011b). Desuden fremkalder mephedron stærkere følelser af trang hos mennesker sammenlignet med MDMA (Brunt et al. 2011), og brugere, der sniffer mephedron, vurderer det som mere vanedannende end kokain (Winstock et al. 2011b). Mephedron indtages af mennesker på en binge-lignende måde (dvs. "stacking") og tages ofte sammen med andre stoffer som cannabis og amfetamin-psykostimulanter (Schifano et al. 2011, Fass et al. 2012, Winstock et al. 2011a, Kelly 2011, Torrance and Cooper 2010). Mephedron findes i stigende grad i tabletter, der sælges som MDMA (Brunt et al. 2011), og brugen af det vil sandsynligvis overgå brugen af MDMA, da renheden af sidstnævnte stof fortsat falder (Brunt et al. 2011, Tanner-Smith 2006, Teng et al. 2006). På baggrund af de almindelige mønstre for misbrug af mephedron og andre "badesalt"-ingredienser er det vigtigt at overveje, om der opstår yderligere sundhedsrisici hos mennesker, når disse stoffer kombineres med amfetamin, bevidst eller ubevidst. Vores resultater, der viser, at mephedron i det mindste øger neurotoksiciteten for DA-nerveender i striatum forårsaget af metamfetamin, amfetamin og MDMA, afslører en særlig farlig og uventet egenskab ved denne β-ketoamfetamin.
Anvendte forkortelser
5-HT serotonin
DA dopamin
DAT DA-transportør
MDMA 3,4-methylendioxymetamfetamin
TH tyrosin hydroxylase
VMAT vesikulær monoamintransportør