La méphédrone (4-méthylméthcathinone) est un stimulant β-cétoamphétamine qui présente d'étroites similitudes structurelles et mécaniques avec la méthamphétamine. L'une des actions les plus puissantes associées à la méphédrone est sa capacité à stimuler la libération de dopamine (DA) et à bloquer sa recapture grâce à son interaction avec le transporteur de la dopamine (DAT). Bien que la méphédrone ne soit pas toxique pour les terminaisons nerveuses DA, sa capacité à bloquer le DAT pourrait offrir une protection contre la neurotoxicité induite par la méthamphétamine, à l'instar d'autres inhibiteurs du DAT. Pour tester cette possibilité, des souris ont été traitées avec de la méphédrone (10, 20 ou 40 mg/kg) avant chaque injection d'un régime neurotoxique de méthamphétamine (4 injections de 2,5 ou 5,0 mg/kg à 2 heures d'intervalle). L'intégrité des terminaisons nerveuses DA du striatum a été évaluée par des mesures des niveaux de DA, de DAT et de tyrosine hydroxylase. La toxicité DA modérée à sévère associée aux différentes doses de méthamphétamine n'a été empêchée par aucune dose de méphédrone et a même été significativement augmentée. L'hyperthermie provoquée par un traitement combiné à la méphédrone et à la méthamphétamine était la même que celle observée après l'administration de l'une ou l'autre de ces substances en monothérapie. La méphédrone a également renforcé les effets neurotoxiques de l'amphétamine et de la MDMA sur les terminaisons nerveuses DA. En revanche, la nomifensine a protégé contre la neurotoxicité induite par la méthamphétamine. Comme la méphédrone augmente la neurotoxicité de la méthamphétamine, les présents résultats suggèrent qu'elle interagit avec le DAT d'une manière différente de celle des autres inhibiteurs typiques du DAT. Les effets relativement inoffensifs de la méphédrone seule sur les terminaisons nerveuses DA masquent une interaction potentiellement dangereuse avec les drogues qui sont souvent consommées en même temps qu'elle, ce qui entraîne une neurotoxicité accrue.
La méphédrone (4-méthylméthcathinone) est un dérivé de la cathinone et un analogue structurel de la méthamphétamine et de la 3,4-méthylènedioxy-méthamphétamine (MDMA). La méphédrone est l'un des ingrédients psychoactifs des "sels de bain", au même titre que d'autres composés tels que la méthylone, la butylone et la 3,4-méthylènedioxypyrovalérone (MDPV). Les β-cétoamphétamines font l'objet d'un abus croissant, en grande partie à cause de la disponibilité très restreinte des précurseurs nécessaires à la synthèse de la méthamphétamine et de la MDMA dans les laboratoires clandestins et de la réduction correspondante de leur pureté (Winstock et al. 2011b, Brunt et al. 2011). L'abus de β-cétoamphétamines ne cessant d'augmenter, la liste de leurs effets indésirables s'est allongée et comprend désormais des complications cardiovasculaires, de l'agitation, de l'insomnie, des psychoses et des dépressions (Schifano et al. 2011, Prosser et Nelson 2012).
En tant que congénères chimiques de la méthamphétamine et de la MDMA, il n'est pas surprenant que les β-cétoamphétamines aient un grand nombre des mêmes effets que ces premières drogues sur le système nerveux central. Par exemple, ces drogues bloquent les transporteurs de la dopamine (DA) et de la sérotonine (5-HT) (DAT et SERT, respectivement) (Cozzi et al. 1999, Rothman et al. 2003, Fleckenstein et al. 2000, Lopez-Arnau et al. 2012) et elles stimulent la libération de monoamines in vitro (Kalix et Glennon 1986, Gygi et al. 1997, Rothman et al. 2003) et in vivo (Gygi et al. 1997, Kehr et al. 2011). La méthcathinone entraîne des réductions persistantes de l'activité de la tryptophane hydroxylase et de la tyrosine hydroxylase (TH) et une déplétion de la DA et de la 5-HT (Gygi et al. 1997, Gygi et al. 1996, Sparago et al. 1996). Des études d'imagerie TEP chez des consommateurs abstinents de méthcathinone ont révélé une réduction de la densité de la DAT dans le striatum, ce qui suggère une perte de terminaux DA (McCann et al. 1998). La stimulation simultanée de la libération de DA et l'inhibition de son absorption reflètent les éléments critiques qui sous-tendent la neurotoxicité associée à la méthamphétamine (Kuhn et al. 2008, Yamamoto et Bankson 2005, Cadet et al. 2007, Fleckenstein et al. 2007).
Nous (Angoa-Perez et al. 2012) et d'autres (Baumann et al. 2012, Hadlock et al. 2011) avons récemment étudié la possibilité que la méphédrone puisse provoquer une neurotoxicité comme la méthamphétamine et la MDMA. De manière surprenante, la méphédrone n'était pas toxique pour les terminaisons nerveuses DA du striatum (Hadlock et al. 2011, Baumann et al. 2012, Angoa-Perez et al. 2012). La question de savoir si la méphédrone endommage les terminaisons nerveuses 5-HT n'est pas tranchée, une étude ayant documenté des effets positifs (Hadlock et al. 2011) et une autre des effets négatifs (Baumann et al. 2012). À la lumière de l'effet relativement bénin de la méphédrone sur les terminaisons nerveuses DA et compte tenu de ses propriétés en tant que bloqueur du DAT, nous avons émis l'hypothèse qu'elle pourrait en fait protéger le système neuronal DA des effets neurotoxiques de la méthamphétamine, comme cela est connu pour se produire avec d'autres bloqueurs du DAT tels que l'acide amphonelique (Pu et al. 1994, Schmidt et Gibb 1985, Marek et al. 1990) et la nomifensine (Poth et al. 2012). Nous rapportons actuellement que la méphédrone augmente de manière significative la neurotoxicité de la méthamphétamine. Cet effet s'étend à l'amphétamine et à la MDMA, des drogues souvent consommées conjointement avec la méphédrone (Feyissa et Kelly 2008, Schifano et al. 2011). Ces résultats surprenants jettent une lumière nouvelle sur l'abus de méphédrone et ajoutent une urgence à la reconnaissance de cette propriété subtile et dangereuse de cette β-cétoamphétamine.
Matériels et méthodes
Médicaments et réactifs
Le chlorhydrate de méphédrone et la 3,4-méthylènedioxyméthamphétamine (MDMA) ont été obtenus auprès du NIDA Research Resources Drug Supply Program. (+) Le chlorhydrate de méthamphétamine, le maléate de nomifensine, le sulfate de d-amphétamine, le pentobarbital, l'AD et tous les tampons et réactifs CLHP ont été achetés à Sigma-Aldrich (St. Louis, MO, USA). Les kits de dosage des protéines de l'acide bicinchoninique ont été obtenus auprès de Pierce (Rockford, IL, USA). Les anticorps polyclonaux contre la TH de rat ont été produits comme décrit précédemment (Kuhn et Billingsley 1987). Les anticorps monoclonaux dirigés contre la DAT de rat ont été généreusement fournis par le Dr Roxanne A. Vaughan (University of North Dakota, Grand Forks, ND, USA). Les anticorps secondaires anti-IgG conjugués à la HRP ont été fournis par Jackson ImmunoResearch Laboratories, Inc. (West Grove, PA, USA).
Animaux
Des souris femelles C57BL/6 (Harlan, Indianapolis, IN, USA) pesant 20-25 g au moment de l'expérimentation ont été hébergées à raison de 5 par cage dans de grandes cages à chaussures dans une pièce éclairée (12 h lumière/obscurité) et à température contrôlée. Des souris femelles ont été utilisées car elles sont connues pour être très sensibles aux dommages neuronaux causés par les amphétamines neurotoxiques et pour maintenir la cohérence avec nos études précédentes sur la neurotoxicité de la méthamphétamine (Thomas et al. 2010, Thomas et al. 2008, Thomas et al. 2009). Les souris avaient libre accès à la nourriture et à l'eau. Le comité institutionnel de soins et d'utilisation de la Wayne State University a approuvé les soins aux animaux et les procédures expérimentales. Toutes les procédures étaient également conformes au NIH Guide for the Care and Use of Laboratory Animals (Guide NIH pour le soin et l'utilisation des animaux de laboratoire).
Procédures pharmacologiques, physiologiques et comportementales
Les souris ont été traitées à la méphédrone selon un schéma de type "binge" comprenant 4 injections de 10, 20 ou 40 mg/kg avec un intervalle de 2 heures entre chaque injection. Lorsqu'il est utilisé pour injecter des amphétamines substituées et des dérivés de la cathinone, ce schéma de traitement en continu entraîne des lésions importantes des terminaisons nerveuses DA. Les doses de méphédrone utilisées actuellement se sont avérées non toxiques pour les terminaisons nerveuses DA (Angoa-Perez et al. 2012). Les souris ont été traitées avec de la méthamphétamine (4X 2,5 ou 5 mg/kg), de l'amphétamine (4X 5 mg/kg) ou de la MDMA (4X 20 mg/kg) seule ou en combinaison avec de la méphédrone. Lorsqu'elles étaient traitées avec deux drogues, les souris recevaient une injection de méphédrone 30 minutes avant chacune des 4 injections de méthamphétamine, d'amphétamine ou de MDMA. Les témoins ont reçu des injections de solution saline physiologique selon le même schéma que celui utilisé pour la méphédrone seule ou en combinaison avec d'autres amphétamines. Pour contrôler les effets d'un inhibiteur du DAT sur la toxicité de la méthamphétamine, les souris ont été traitées avec de la nomifensine (4X 5 mg/kg) 30 minutes avant chaque injection de méthamphétamine (4X 5 mg/kg). Toutes les injections ont été réalisées par voie i.p. Les souris ont été sacrifiées 2 jours après le dernier traitement médicamenteux, lorsque la neurotoxicité associée aux amphétamines a atteint son maximum. La température corporelle a été surveillée par télémétrie à l'aide de transpondeurs de température implantables IPTT-300 de Bio Medic Data Systems, Inc. (Seaford, DE, USA). Les températures ont été enregistrées de manière non invasive toutes les 20 minutes, en commençant 60 minutes avant la première injection de METH et en continuant pendant 9 heures à l'aide du système de console DAS-5001 de Bio Medic.
Détermination de la teneur en DA du striatum
Le tissu striatal a été disséqué bilatéralement du cerveau après le traitement et stocké à -80°C. Les tissus congelés ont été pesés et soniqués dans 10 volumes d'acide perchlorique 0,16 N à 4°C. Les protéines insolubles ont été éliminées par centrifugation. Les protéines insolubles ont été éliminées par centrifugation et la DA a été déterminée par HPLC avec détection électrochimique comme décrit précédemment pour la méthamphétamine (Thomas et al. 2010, Thomas et al, 2009).
Détermination des niveaux de protéines TH et DAT par immunoblotting
Les effets des traitements médicamenteux sur les niveaux de TH et de DAT striataux ont été déterminés par immunoblotting comme indice de toxicité pour les terminaisons nerveuses DA striatales. Les souris ont été sacrifiées par décapitation après le traitement et le striatum a été disséqué bilatéralement. Les tissus ont été conservés à -80°C. Le tissu congelé a été désagrégé par sonication dans 1% de SDS à 95°C et le matériel insoluble a été sédimenté par centrifugation. Les protéines ont été déterminées par la méthode de l'acide bicinchoninique et des quantités égales de protéines (70 μg/lane) ont été résolues par électrophorèse sur gel de SDS-polyacrylamide, puis électrofilmées sur nitrocellulose. Les taches ont été bloquées dans une solution saline tamponnée au Tris contenant du Tween 20 (0,1 % v/v) et du lait sec non gras à 5 % pendant 1 h à température ambiante. Des anticorps primaires contre la TH (1:1000) ou la DAT (1:1000) ont été ajoutés aux blots et laissés à incuber pendant 16 heures à 4°C. Les blots ont été lavés 3 fois dans une solution saline tamponnée au Tris pour éliminer les anticorps n'ayant pas réagi, puis incubés avec un anticorps secondaire anti-IgG conjugué à la HRP (1:4000) pendant 1 h à température ambiante. Les bandes immunoréactives ont été visualisées par chimiluminescence renforcée et les densités relatives des bandes réactives à la TH et à la DAT ont été déterminées par imagerie avec un Kodak Image Station (Carestream Molecular Systems, Rochester, NY, USA) et quantifiées à l'aide du logiciel ImageJ (NIH).
Analyse des données
Des ANOVA à deux voies ont été réalisées pour analyser les effets de la méthamphétamine et de la méphédrone sur la DA, la DAT et la TH. Les effets des traitements médicamenteux sur la teneur en DA, TH et DAT du striatum ont fait l'objet d'un test de signification par ANOVA à sens unique suivi du test de comparaison multiple de Tukey. Les résultats des traitements médicamenteux sur la température corporelle centrale au fil du temps ont été analysés à l'aide d'une ANOVA à deux voies suivie d'un test de Bonferroni pour déterminer la signification des différences de température à des moments individuels après le traitement. Les différences ont été considérées comme significatives si p < 0,05. Toutes les analyses statistiques ont été effectuées à l'aide de GraphPad Prism version 5.02 pour Windows (GraphPad Software, San Diego, CA, USA, www.graphpad.com).
Aller à :
Résultats
Effets de la méphédrone sur la neurotoxicité induite par la méthamphétamine
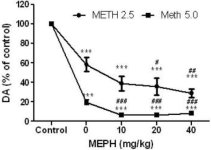
La méphédrone, à des doses (10, 20 ou 40 mg/kg) connues pour ne pas provoquer de toxicité des terminaisons nerveuses DA (Angoa-Perez et al. 2012), a été administrée 30 minutes avant chaque injection de méthamphétamine. La méthamphétamine a été administrée à des doses qui causent des dommages modérés (4X 2,5 mg/kg) ou sévères (4X 5 mg/kg) aux terminaisons nerveuses DA du striatum (Thomas et al. 2004, Thomas et al. 2010). Les résultats présentés dans la figure 1 montrent que les effets principaux de la dose de méthamphétamine (F1,40 = 66,60, p < 0,0001) et de la dose de méphédrone (F4,40 = 131,3, p < 0,0001) sur les niveaux de DA dans le striatum étaient hautement significatifs selon l'ANOVA à deux voies. L'effet principal de la méphédrone administrée en combinaison avec 2,5 mg/kg (F4,22 = 35,96, p < 0,001) ou 5,0 mg/kg de méthamphétamine (F4,17 = 953,9, p < 0,0001) était également très significatif selon l'ANOVA à sens unique. Tous les traitements avec l'une ou l'autre dose de méthamphétamine ± méphédrone ont entraîné des réductions significativement plus importantes de la DA par rapport au contrôle respectif (p < 0,0001 pour tous). La figure 1 montre également que des doses de méphédrone de 20 (p < 0,01) et 40 mg/kg (p < 0,001) ont renforcé de manière significative les effets d'épuisement de 2,5 mg/kg de méthamphétamine sur l'AD, tandis que toutes les doses de méphédrone ont renforcé de manière significative les effets de 5,0 mg/kg de méthamphétamine sur les niveaux d'AD (p < 0,0001 pour toutes les doses).
Fig.1 : Effets de la méphédrone sur la méthamphétamine
Effets de la méphédrone sur les réductions de l'AD striatale induites par la méthamphétamine. Les souris ont été traitées avec les doses indiquées de méphédrone (MEPH) 30 minutes avant chaque injection de 2,5 (-) ou 5,0 mg/kg (■) de méthamphétamine (METH) et ont été sacrifiées 2 jours plus tard pour la détermination des niveaux striataux de DA par HPLC. Les données sont des moyennes ± SEM pour 5-7 souris par groupe. Certaines barres d'erreur étaient trop petites pour dépasser la taille des symboles et ne sont pas visibles. ***p < 0,001 par rapport aux témoins et #p < 0,01, ##p < 0,001 ou ###p < 0,0001 par rapport à la dose respective de méthamphétamine (test de comparaison multiple de Tukey).
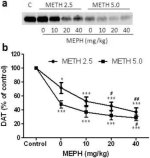
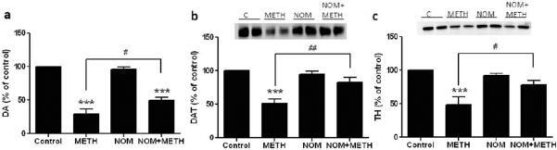
La figure 2a montre que la méphédrone a augmenté de manière significative les réductions des niveaux de DAT induites par la méthamphétamine, comme l'a montré l'immunoblot. Les immunoblots ont été quantifiés et, conformément aux résultats obtenus pour la DA, les effets principaux de la dose de méthamphétamine (F1,92 = 9,48, p < 0,001) et de la dose de méphédrone (F4,92 = 37,56, p < 0,0001) sur les taux de DAT dans le striatum ont été hautement significatifs d'après l'ANOVA à deux voies (Fig. 2b). L'effet principal de la méphédrone administrée en association avec 2,5 mg/kg (F4,56 = 15,55, p < 0,0001) ou 5,0 mg/kg de méthamphétamine (F4,39 = 24,84, p < 0,0001) était également très significatif selon l'ANOVA à sens unique. Tous les traitements avec l'une ou l'autre dose de méthamphétamine ± méphédrone ont entraîné des réductions significativement plus importantes de la DAT par rapport au contrôle respectif (p < 0,01 pour 2,5 mg/kg de méthamphétamine seule ; p < 0,0001 pour tous les autres traitements). La figure 2b montre également que les doses de méphédrone de 20 mg/kg (p < 0,01) et de 40 mg/kg (p < 0,001) ont renforcé de manière significative les réductions de DAT provoquées par 2,5 mg/kg de méthamphétamine, alors que seule la dose de 40 mg/kg de méphédrone a renforcé de manière significative (p < 0,01) les effets de 5,0 mg/kg de méthamphétamine sur les réductions de DAT.
Fig.2
Effets de la méphédrone sur les réductions de la DAT striatale induites par la méthamphétamine. Les souris ont été traitées avec les doses indiquées de méphédrone (MEPH) 30 min avant chaque injection de 2,5 (●) ou 5,0 mg/kg (■) de méthamphétamine (METH) et sacrifiées 2j plus tard pour la détermination des niveaux striataux de DAT par immunoblotting (a). Les taches ont été quantifiées à l'aide d'ImageJ et les données sont des moyennes ± SEM pour 10-12 souris par groupe (b). *p < 0,01 ou ***p < 0,0001 vs contrôle (C) et #p < 0,01 ou ##p < 0,001 vs la dose respective de méthamphétamine (test de comparaison multiple de Tukey).
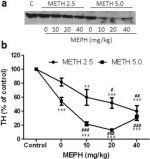
La figure 3a montre que la méphédrone a augmenté de manière significative les réductions des niveaux de TH induites par la méthamphétamine, comme l'a montré l'immunoblot. Les immunoblots ont été quantifiés et, conformément aux résultats ci-dessus pour la DA et la DAT, les effets principaux de la dose de méthamphétamine (F1,81 = 47,89, p < 0,0001) et de la dose de méphédrone (F4,81 = 63,57, p < 0,0001) ont été hautement significatifs par ANOVA à deux voies (Fig. 3b). L'effet principal de la méphédrone administrée en combinaison avec 2,5 mg/kg (F4,34 = 12,98, p < 0,0001) ou 5,0 mg/kg de méthamphétamine (F4,49 = 99,16, p < 0,0001) était également très significatif selon l'ANOVA à une voie. Tous les traitements avec l'une ou l'autre dose de méthamphétamine ± méphédrone ont entraîné des réductions significativement plus importantes du TH par rapport au contrôle respectif (p < 0,001 pour 2,5 mg/kg de méthamphétamine + 10 mg/kg de méphédrone ; p < 0,0001 pour toutes les autres combinaisons), à l'exception de 2,5 mg/kg de méthamphétamine seule qui n'a pas modifié de manière significative les niveaux de TH (c.-à-d. pas de toxicité). La figure 3b montre également que les doses de méphédrone de 20 mg/kg (p < 0,01) et de 40 mg/kg (p < 0,001) ont renforcé de manière significative les réductions de TH provoquées par 2,5 mg/kg de méthamphétamine et que les trois doses de méphédrone ont renforcé de manière significative (p < 0,0001) les effets de 5,0 mg/kg de méthamphétamine sur les réductions de TH.
Fig. 3
Effets de la méphédrone sur les réductions de la TH striatale induites par la méthamphétamine. Les souris ont été traitées avec les doses indiquées de méphédrone (MEPH) 30 min avant chaque injection de 2,5 (●) ou 5,0 mg/kg (■) de méthamphétamine (METH) et sacrifiées 2j plus tard pour la détermination des niveaux striataux de TH par immunoblotting (a). Les blots ont été quantifiés à l'aide d'ImageJ et les données sont des moyennes ± SEM pour 10-12 souris par groupe (b). Certaines barres d'erreur étaient trop petites pour dépasser la taille des symboles et ne sont pas visibles. **p < 0,001 ou ***p < 0,0001 vs contrôle (C) et #p < 0,01, ##p < 0,001 ou ###p < 0,0001) vs la dose respective de méthamphétamine (test de comparaison multiple de Tukey).
Effets de la méphédrone sur l'hyperthermie induite par la méthamphétamine
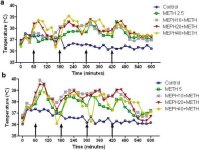
La méphédrone, comme la méthamphétamine, provoque une hyperthermie importante (Hadlock et al. 2011, Baumann et al. 2012, Angoa-Perez et al. 2012). Lorsque la méphédrone a été administrée 30 minutes avant chaque injection de méthamphétamine, la figure 4 montre que les effets principaux des doses de méthamphétamine et de méphédrone (F1,300 = 11,99, p < 0,0001) sur le temps (F4,300 = 51,73, p < 0,0001) sont très significatifs selon l'ANOVA à deux voies. Les effets principaux de la méphédrone administrée en association avec 2,5 mg/kg de méthamphétamine (F4,120 = 41,44, p < 0,0001, panel a) dans le temps (F30,120 = 3,84, p < 0,0001) ou 5,0 mg/kg de méthamphétamine (F4,120 = 78,09, p < 0,0001, panel b) dans le temps (F30,120 = 9,98, p < 0,0001) ont également été hautement significatifs par ANOVA à deux voies. Tous les traitements avec l'une ou l'autre dose de méthamphétamine ± méphédrone étaient significativement différents des contrôles respectifs (p < 0,0001 pour tous les traitements).
Fig. 4
Effets de la méphédrone sur l'hyperthermie induite par la méthamphétamine. Les souris ont été traitées avec les doses indiquées de méphédrone (MEPH) 30 minutes avant chaque injection de 2,5 (a) ou 5,0 mg/kg (b) de méthamphétamine (METH). Les températures centrales ont été mesurées à intervalles de 20 minutes par télémétrie à partir de 60 minutes avant la première injection de méthamphétamine. Les 4 injections de méthamphétamine sont indiquées par les flèches reposant sur l'axe des x. Les données sont exprimées en tant que température corporelle moyenne de 6-8 souris par groupe. Les SEM sont toujours inférieures à 10 % de la moyenne et sont omises par souci de clarté.
Effets de la méphédrone sur la neurotoxicité induite par l'amphétamine et la MDMA
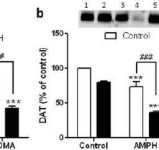
Afin de vérifier si les effets de la méphédrone sur la méthamphétamine pouvaient être étendus à d'autres amphétamines neurotoxiques, des souris ont été traitées avec cette β-cétoamphétamine (20 mg/kg) et de l'amphétamine (4X 5 mg/kg) ou de la MDMA (4X 20 mg/kg), et les résultats sont présentés dans la figure 5. Rappelons que la méphédrone elle-même ne réduit pas la DA, la DAT ou la TH striatale (Angoa-Perez et al. 2012). L'effet principal de la drogue (F5,27 = 27,18, p < 0,0001) était hautement significatif par ANOVA à sens unique pour les réductions de DA (Fig. 5a). La figure 5a montre également que tous les traitements à l'amphétamine (p < 0,001) ou à la MDMA (p < 0,001) seuls ou en combinaison avec la méphédrone (p < 0,0001 pour les deux drogues) ont réduit de manière significative les niveaux de DA par rapport au contrôle. La méphédrone a renforcé de manière significative les réductions de DA provoquées par l'amphétamine (p < 0,01) ou la MDMA (p < 0,01). La figure 5b montre des effets similaires des traitements combinés sur les niveaux de DAT dans le striatum. L'effet principal de la drogue (F4,49 = 42,63, p < 0,0001) était très significatif dans l'ANOVA à sens unique pour la DAT. La figure 5b montre également que tous les traitements à l'amphétamine ou à la MDMA étaient significativement (p < 0,0001 pour tous) inférieurs au contrôle. La méphédrone a également renforcé de manière significative les réductions de DAT causées par l'amphétamine ou la MDMA (p < 0,0001 dans les deux cas). Enfin, la figure 5c montre que l'effet principal de la drogue (F4,50 = 75,06, p < 0,0001) était très significatif dans l'ANOVA à sens unique pour les réductions de TH. La figure 5c montre également que tous les traitements à l'amphétamine ou à la MDMA étaient significativement (p < 0,0001 pour tous) inférieurs au contrôle. La méphédrone a également renforcé de manière significative les réductions de TH causées par l'amphétamine ou la MDMA (p < 0,0001 dans les deux cas).
Fig. 5
Effets de la méphédrone sur la neurotoxicité des terminaisons nerveuses DA induite par l'amphétamine ou la MDMA. Des souris ont été traitées avec 20 mg/kg de méphédrone (MEPH) 30 minutes avant chaque injection de 5,0 mg/kg d'amphétamine (AMPH) ou de 20 mg/kg de MDMA et ont été sacrifiées 2 jours après le traitement pour la détermination des niveaux striataux de (a) DA par HPLC. (b) DAT et (c) TH ont été déterminés par immunoblotting et les blots ont été quantifiés à l'aide d'ImageJ. Les immunoblots représentatifs de la DAT et de la TH sont inclus dans les encadrés des panneaux (b) et (c) respectivement et les traitements pour les deux panneaux sont indiqués par 1,5 : contrôle ; 2,6 : MEPH ; 3 : AMPH ; 4 : AMPH + MEPH ; 7 : MDMA ; et 8 : MDMA + MEPH. Les données sont des moyennes ± SEM pour 5-12 souris dans chaque groupe. **p < 0,001 ou ***p < 0,0001 vs contrôle et #p < 0,01 ou ###p < 0,0001 vs AMPH ou MDMA (test de comparaison multiple de Tukey).
Effets de la nomifensine sur la neurotoxicité induite par la méthamphétamine
La nomifensine, un puissant bloqueur du DAT sans potentiel connu d'abus ou de neurotoxicité, a été testée pour sa capacité à protéger contre la neurotoxicité induite par la méthamphétamine et pour contraster avec les actions de la méphédrone sur la toxicité des terminaisons nerveuses DA causée par la méthamphétamine, l'amphétamine et la MDMA. Les résultats de la figure 6a montrent que l'effet principal de la drogue (F3,16 = 63,39, p < 0,0001) sur les niveaux de DA était très significatif selon l'ANOVA à sens unique. La nomifensine seule n'a pas modifié les niveaux de DA, mais la réduction causée par la méthamphétamine (p < 0,0001) a été légèrement mais significativement inversée par la nomifensine (p < 0,01). L'effet principal de la drogue (F3,20 = 16,78, p < 0,0001) sur les niveaux de DAT était très significatif selon l'ANOVA à sens unique, comme le montre la figure 6b. La nomifensine n'a pas modifié les niveaux de DAT mais a fourni une protection significative (p < 0,001) contre la réduction de la DAT striatale causée par la méthamphétamine (p < 0,0001) par rapport au contrôle. Enfin, la figure 6c montre que l'effet principal de la drogue (F3,15 = 14,10, p < 0,0001) sur les niveaux de TH était très significatif selon l'ANOVA à sens unique. Comme pour la DA et la DAT, la réduction de la TH causée par la méthamphétamine (p < 0,0001) a été légèrement mais significativement empêchée par la nomifensine (p < 0,01).
Fig. 6
Effets de la nomifensine sur la neurotoxicité des terminaisons nerveuses DA induite par la méthamphétamine. Des souris ont été traitées avec 5,0 mg/kg de nomifensine (NOM) 30 minutes avant chaque injection de 5,0 mg/kg de méthamphétamine (METH) et ont été sacrifiées deux jours plus tard pour la détermination des niveaux striataux de (a) DA par HPLC. (b) DAT et (c) TH ont été déterminés par immunoblotting et les blots ont été quantifiés à l'aide d'ImageJ. Des immunoblots représentatifs de la DAT et de la TH sont inclus dans les encadrés des panneaux (b) et (c) respectivement. Les données sont des moyennes plus SEM pour 5-7 souris par groupe. ***p < 0,0001 vs contrôle (C) et #p < 0,01 ou ##p < 0,001 vs méthamphétamine seule (test de comparaison multiple de Tukey).
Discussion
L'objectif de la présente étude était de déterminer si la méphédrone pouvait prévenir la toxicité des terminaisons nerveuses DA causée par la méthamphétamine. Sur la base de sa similarité chimique avec la méthamphétamine et la MDMA, on s'attendait initialement à ce que la méphédrone exerce des effets nocifs sur le système neuronal DA. Cependant, plusieurs études ont établi presque simultanément que la méphédrone n'était pas toxique pour les terminaisons nerveuses DA (Angoa-Perez et al. 2012, Baumann et al. 2012, Hadlock et al. 2011). La question de savoir si ce médicament cause des dommages au système neuronal 5-HT reste ouverte. Une étude a fait état de réductions persistantes de la fonction des terminaisons nerveuses 5-HT (Hadlock et al. 2011), tandis qu'une autre a montré que la méphédrone ne causait pas de dommages (Baumann et al. 2012). La méphédrone interagit avec la terminaison nerveuse DA d'une manière qui suggère qu'elle stimule effectivement la libération et bloque la recapture de la DA via ses interactions avec le DAT. Un aspect clé du mécanisme d'action neurotoxique de la méthamphétamine est sa capacité à accéder aux terminaisons nerveuses DA par l'intermédiaire du DAT et à perturber l'homéostasie de la DA (Sulzer 2011). Si cette étape précoce de la cascade neurotoxique de la méthamphétamine est empêchée par l'inhibition du DAT, la toxicité est évitée (Pu et al. 1994, Poth et al. 2012, Marek et al. 1990, Schmidt et Gibb 1985). Nous avons pensé que la méphédrone pourrait avoir la même propriété protectrice que d'autres inhibiteurs de la DAT, mais nous avons observé au contraire une augmentation significative de la toxicité. Cette interaction a été observée en utilisant deux doses différentes de méthamphétamine qui causent des dommages modérés ou sévères aux terminaisons nerveuses DA (4X 2,5 ou 5,0 mg/kg, respectivement). Cet effet potentialisateur de la méphédrone ne s'est pas limité à la méthamphétamine et s'est étendu à l'amphétamine et à la MDMA, deux drogues souvent consommées conjointement avec la méphédrone et d'autres β-cétoamphétamines (Feyissa et Kelly 2008, Schifano et al. 2011, Kelly 2011). Par conséquent, bien que la méphédrone ne soit pas toxique pour les terminaisons nerveuses DA du striatum, elle potentialise les effets neurotoxiques d'autres drogues d'abus. Cette nouvelle découverte devrait faire apparaître l'abus de méphédrone sous un jour encore plus sombre, car son absence de neurotoxicité intrinsèque peut le faire paraître inoffensif.
L'hyperthermie est un effet indésirable aigu couramment signalé de l'ingestion de méthamphétamine (Greene et al. 2008) et de β-cétoamphétamine chez l'homme (Borek et Holstege 2012, Prosser et Nelson 2012). Comme la méthamphétamine, de nombreuses drogues de type β-cétoamphétamine provoquent également des élévations significatives de la température centrale chez les rongeurs (Angoa-Perez et al. 2012, Hadlock et al. 2011, Baumann et al. 2012, Rockhold et al. 1997). Si l'hyperthermie provoquée par la méthamphétamine peut contribuer à ses effets morphologiques et neuronaux, elle n'est pas nécessairement la cause directe de ces effets (Kiyatkin et Sharma 2009). Nous avons enregistré la température corporelle centrale de souris traitées à la méphédrone et à la méthamphétamine et avons observé que le traitement combiné n'augmentait pas la température au-delà des augmentations maximales observées après l'administration de l'une ou l'autre des drogues seules. La méthamphétamine a provoqué une augmentation de la température corporelle liée à la dose et cette hyperthermie était invariable sur l'ensemble de la gamme de doses de méphédrone testée. En fait, la baisse de la température corporelle observée après l'injection de méphédrone (Angoa-Perez et al. 2012) a été maintenue à des doses plus élevées de méphédrone et de méthamphétamine. Même si l'hyperthermie induite par la drogue n'a pas été renforcée par un traitement combiné, les effets neurotoxiques se sont additionnés. Par conséquent, au moins dans le cas présent, il semble que les effets neurotoxiques de la méthamphétamine peuvent être renforcés par la méphédrone d'une manière indépendante de l'hyperthermie.
La méphédrone inhibe clairement la fonction du DAT et bloque la recapture de la DA in vitro (Lopez-Arnau et al. 2012, Hadlock et al. 2011, Kehr et al. 2011, Martinez-Clemente et al. 2012, Cozzi et al. 1999). La méphédrone déplace le WIN-35,428 de son site de liaison sur le DAT, ce qui suggère qu'il s'agit d'un inhibiteur compétitif de l'absorption de DA (Martinez-Clemente et al. 2012, Lopez-Arnau et al. 2012). La puissance de la méphédrone à cet égard est très similaire à celle de la méthamphétamine (Cozzi et al. 1999) et de la MDMA (Escubedo et al. 2011). On ne sait pas si la méphédrone est transportée par le DAT, mais la méthcathinone l'est (Cozzi et Foley 2003). La nomifensine et l'acide amphonelique, qui se lient au DAT et inhibent le captage de la DA, offrent une protection substantielle contre la neurotoxicité induite par la méthamphétamine (Pu et al. 1994, Marek et al. 1990, Schmidt et Gibb 1985, Poth et al. 2012) et les souris dépourvues de DAT sont résistantes à la toxicité neuronale de la méthamphétamine (Fumagalli et al. 1998). Sachant que la méphédrone n'est pas neurotoxique et qu'elle est un bloqueur du DAT, on peut prédire qu'elle devrait prévenir la toxicité. Nous avons testé la nomifensine à cet égard en tant que contrôle positif et avons confirmé qu'elle protège contre la déplétion de DA, DAT et TH induite par la méthamphétamine. La nomifensine inhibe également le transporteur de la norépinéphrine (Brogden et al. 1979), mais cette propriété ne peut pas expliquer les présents résultats, car la plupart des β-cétoamphétamines, y compris la méphédrone, inhibent le transporteur de la norépinéphrine et bloquent l'absorption de la norépinéphrine (Kelly 2011, Rothman et al. 2003, Cozzi et al. 1999, Sogawa et al. 2011, Lopez-Arnau et al. 2012). Un rôle du système neuronal 5-HT dans certaines des actions pharmacologiques de la méphédrone est possible à la lumière de la capacité de cette drogue, comme la MDMA (Yamamoto et al. 1995), à provoquer un efflux de DA striatal via ses interactions avec les récepteurs 5-HT2A (Lopez-Arnau et al. 2012, Martinez-Clemente et al. 2012). L'hyper-locomotion provoquée par la méphédrone est dépendante de la 5-HT endogène (Lopez-Arnau et al. 2012) et cette drogue stimule également la libération de 5-HT et inhibe sa captation in vitro (Sogawa et al. 2011, Cozzi et al. 1999, Nagai et al. 2007, Hadlock et al. 2011, Lopez-Arnau et al. 2012, Martinez-Clemente et al. 2012) et in vivo (Baumann et al. 2012, Kehr et al. 2011). Cependant, nous pouvons exclure un rôle pour la 5-HT endogène dans la neurotoxicité DA, au moins de la méthamphétamine, en montrant que les souris génétiquement dépourvues de 5-HT conservent leur sensibilité à la neurotoxicité (Thomas et al. 2010).
La méphédrone pourrait renforcer la neurotoxicité de la méthamphétamine par plusieurs mécanismes possibles. Tout d'abord, la méphédrone pourrait interagir avec le VMAT pour provoquer une fuite de DA dans le cytoplasme de la terminaison nerveuse présynaptique. Les traitements qui augmentent le pool cytoplasmique (c.-à-d. libérable par la drogue) de DA augmentent la neurotoxicité de la méthamphétamine (Thomas et al. 2008, Thomas et al. 2009, Schmidt et al. 1985). Ce mécanisme n'est pas probable car la méthcathinone n'interagit que faiblement avec le VMAT (Cozzi et al. 1999). Deuxièmement, l'association de la méphédrone et de la méthamphétamine pourrait avoir un effet synergique sur la libération non vésiculaire de DA, mais cette possibilité semble également peu probable à la lumière des résultats montrant que le traitement de cellules CHO exprimant le DAT ou le SERT avec de la méthylone et de la méthamphétamine n'a pas d'effet additif sur la libération de DA ou de 5-HT (Sogawa et al. 2011). Troisièmement, la méphédrone pourrait interagir avec le DAT d'une nouvelle manière qui contribue à la toxicité additive. Il a été démontré que la méthylone associée à la méthamphétamine provoque une cytotoxicité synergique dans les cellules CHO exprimant le DAT ou le SERT, mais pas dans les cellules CHO de type sauvage dépourvues de ces transporteurs (Sogawa et al. 2011). La cytotoxicité observée dans les cellules cultivées dans ces études (c'est-à-dire la libération de LDH) est très différente des dommages causés aux terminaisons nerveuses DA par la méthamphétamine, mais ce mécanisme suggère un rôle intéressant mais non défini pour le DAT dans la cytotoxicité accrue. Enfin, la méphédrone pourrait modifier le métabolisme de la méthamphétamine. La méphédrone est principalement métabolisée par N-déméthylation (Meyer et Maurer 2010), tout comme la méthamphétamine et la MDMA (Caldwell 1976). Ce mécanisme est étayé par la démonstration que la méthamphétamine et la MDMA inhibent mutuellement la production de leurs métabolites primaires respectifs et élèvent les concentrations plasmatiques de la drogue au-dessus de celles observées après l'administration de l'une ou l'autre drogue seule (Kuwayama et al. 2012). Les doses de méphédrone utilisées actuellement et dans notre étude précédente (Angoa-Perez et al. 2012), bien qu'élevées, ne sont pas neurotoxiques et se situent dans la gamme des drogues consommées par les humains (McErath et O'Neill 2011). Par conséquent, la méphédrone pourrait agir comme la MDMA pour augmenter les taux plasmatiques de méthamphétamine en inhibant son métabolisme. Une analyse pharmacocinétique approfondie sera nécessaire pour confirmer cette dernière possibilité.
L'abus des β-cétoamphétamines augmente à un rythme alarmant et la méphédrone est désormais l'une des drogues les plus couramment consommées après le cannabis, la MDMA et la cocaïne (Morris 2010, Winstock et al. 2011b). En outre, la méphédrone induit chez l'homme des sensations de manque plus fortes que la MDMA (Brunt et al. 2011) et les usagers qui sniffent la méphédrone la considèrent comme plus addictive que la cocaïne (Winstock et al. 2011b). La méphédrone est consommée par les humains sous forme de "binge" (c'est-à-dire de "stacking") et est souvent prise avec d'autres drogues telles que le cannabis et les psychostimulants amphétaminiques (Schifano et al. 2011, Fass et al. 2012, Winstock et al. 2011a, Kelly 2011, Torrance et Cooper 2010). La méphédrone se trouve de plus en plus souvent dans des comprimés vendus comme de la MDMA (Brunt et al. 2011) et sa consommation dépassera probablement celle de la MDMA, car la pureté de cette dernière drogue continue de baisser (Brunt et al. 2011, Tanner-Smith 2006, Teng et al. 2006). Compte tenu des schémas courants d'abus de méphédrone et d'autres ingrédients des "sels de bain", il est important de se demander si des risques supplémentaires pour la santé humaine apparaissent lorsque ces drogues sont combinées volontairement ou involontairement avec des amphétamines. Nos résultats montrant qu'au moins la méphédrone renforce de manière significative la neurotoxicité des terminaisons nerveuses DA du striatum causée par la méthamphétamine, l'amphétamine et la MDMA révèlent une propriété particulièrement dangereuse et inattendue de cette β-cétoamphétamine.
Abréviations utilisées
5-HT sérotonine
DA dopamine
DAT transporteur DA
MDMA 3,4-méthylènedioxyméthamphétamine
TH tyrosine hydroxylase
VMAT transporteur vésiculaire de monoamines
La méphédrone (4-méthylméthcathinone) est un dérivé de la cathinone et un analogue structurel de la méthamphétamine et de la 3,4-méthylènedioxy-méthamphétamine (MDMA). La méphédrone est l'un des ingrédients psychoactifs des "sels de bain", au même titre que d'autres composés tels que la méthylone, la butylone et la 3,4-méthylènedioxypyrovalérone (MDPV). Les β-cétoamphétamines font l'objet d'un abus croissant, en grande partie à cause de la disponibilité très restreinte des précurseurs nécessaires à la synthèse de la méthamphétamine et de la MDMA dans les laboratoires clandestins et de la réduction correspondante de leur pureté (Winstock et al. 2011b, Brunt et al. 2011). L'abus de β-cétoamphétamines ne cessant d'augmenter, la liste de leurs effets indésirables s'est allongée et comprend désormais des complications cardiovasculaires, de l'agitation, de l'insomnie, des psychoses et des dépressions (Schifano et al. 2011, Prosser et Nelson 2012).
En tant que congénères chimiques de la méthamphétamine et de la MDMA, il n'est pas surprenant que les β-cétoamphétamines aient un grand nombre des mêmes effets que ces premières drogues sur le système nerveux central. Par exemple, ces drogues bloquent les transporteurs de la dopamine (DA) et de la sérotonine (5-HT) (DAT et SERT, respectivement) (Cozzi et al. 1999, Rothman et al. 2003, Fleckenstein et al. 2000, Lopez-Arnau et al. 2012) et elles stimulent la libération de monoamines in vitro (Kalix et Glennon 1986, Gygi et al. 1997, Rothman et al. 2003) et in vivo (Gygi et al. 1997, Kehr et al. 2011). La méthcathinone entraîne des réductions persistantes de l'activité de la tryptophane hydroxylase et de la tyrosine hydroxylase (TH) et une déplétion de la DA et de la 5-HT (Gygi et al. 1997, Gygi et al. 1996, Sparago et al. 1996). Des études d'imagerie TEP chez des consommateurs abstinents de méthcathinone ont révélé une réduction de la densité de la DAT dans le striatum, ce qui suggère une perte de terminaux DA (McCann et al. 1998). La stimulation simultanée de la libération de DA et l'inhibition de son absorption reflètent les éléments critiques qui sous-tendent la neurotoxicité associée à la méthamphétamine (Kuhn et al. 2008, Yamamoto et Bankson 2005, Cadet et al. 2007, Fleckenstein et al. 2007).
Nous (Angoa-Perez et al. 2012) et d'autres (Baumann et al. 2012, Hadlock et al. 2011) avons récemment étudié la possibilité que la méphédrone puisse provoquer une neurotoxicité comme la méthamphétamine et la MDMA. De manière surprenante, la méphédrone n'était pas toxique pour les terminaisons nerveuses DA du striatum (Hadlock et al. 2011, Baumann et al. 2012, Angoa-Perez et al. 2012). La question de savoir si la méphédrone endommage les terminaisons nerveuses 5-HT n'est pas tranchée, une étude ayant documenté des effets positifs (Hadlock et al. 2011) et une autre des effets négatifs (Baumann et al. 2012). À la lumière de l'effet relativement bénin de la méphédrone sur les terminaisons nerveuses DA et compte tenu de ses propriétés en tant que bloqueur du DAT, nous avons émis l'hypothèse qu'elle pourrait en fait protéger le système neuronal DA des effets neurotoxiques de la méthamphétamine, comme cela est connu pour se produire avec d'autres bloqueurs du DAT tels que l'acide amphonelique (Pu et al. 1994, Schmidt et Gibb 1985, Marek et al. 1990) et la nomifensine (Poth et al. 2012). Nous rapportons actuellement que la méphédrone augmente de manière significative la neurotoxicité de la méthamphétamine. Cet effet s'étend à l'amphétamine et à la MDMA, des drogues souvent consommées conjointement avec la méphédrone (Feyissa et Kelly 2008, Schifano et al. 2011). Ces résultats surprenants jettent une lumière nouvelle sur l'abus de méphédrone et ajoutent une urgence à la reconnaissance de cette propriété subtile et dangereuse de cette β-cétoamphétamine.
Matériels et méthodes
Médicaments et réactifs
Le chlorhydrate de méphédrone et la 3,4-méthylènedioxyméthamphétamine (MDMA) ont été obtenus auprès du NIDA Research Resources Drug Supply Program. (+) Le chlorhydrate de méthamphétamine, le maléate de nomifensine, le sulfate de d-amphétamine, le pentobarbital, l'AD et tous les tampons et réactifs CLHP ont été achetés à Sigma-Aldrich (St. Louis, MO, USA). Les kits de dosage des protéines de l'acide bicinchoninique ont été obtenus auprès de Pierce (Rockford, IL, USA). Les anticorps polyclonaux contre la TH de rat ont été produits comme décrit précédemment (Kuhn et Billingsley 1987). Les anticorps monoclonaux dirigés contre la DAT de rat ont été généreusement fournis par le Dr Roxanne A. Vaughan (University of North Dakota, Grand Forks, ND, USA). Les anticorps secondaires anti-IgG conjugués à la HRP ont été fournis par Jackson ImmunoResearch Laboratories, Inc. (West Grove, PA, USA).
Animaux
Des souris femelles C57BL/6 (Harlan, Indianapolis, IN, USA) pesant 20-25 g au moment de l'expérimentation ont été hébergées à raison de 5 par cage dans de grandes cages à chaussures dans une pièce éclairée (12 h lumière/obscurité) et à température contrôlée. Des souris femelles ont été utilisées car elles sont connues pour être très sensibles aux dommages neuronaux causés par les amphétamines neurotoxiques et pour maintenir la cohérence avec nos études précédentes sur la neurotoxicité de la méthamphétamine (Thomas et al. 2010, Thomas et al. 2008, Thomas et al. 2009). Les souris avaient libre accès à la nourriture et à l'eau. Le comité institutionnel de soins et d'utilisation de la Wayne State University a approuvé les soins aux animaux et les procédures expérimentales. Toutes les procédures étaient également conformes au NIH Guide for the Care and Use of Laboratory Animals (Guide NIH pour le soin et l'utilisation des animaux de laboratoire).
Procédures pharmacologiques, physiologiques et comportementales
Les souris ont été traitées à la méphédrone selon un schéma de type "binge" comprenant 4 injections de 10, 20 ou 40 mg/kg avec un intervalle de 2 heures entre chaque injection. Lorsqu'il est utilisé pour injecter des amphétamines substituées et des dérivés de la cathinone, ce schéma de traitement en continu entraîne des lésions importantes des terminaisons nerveuses DA. Les doses de méphédrone utilisées actuellement se sont avérées non toxiques pour les terminaisons nerveuses DA (Angoa-Perez et al. 2012). Les souris ont été traitées avec de la méthamphétamine (4X 2,5 ou 5 mg/kg), de l'amphétamine (4X 5 mg/kg) ou de la MDMA (4X 20 mg/kg) seule ou en combinaison avec de la méphédrone. Lorsqu'elles étaient traitées avec deux drogues, les souris recevaient une injection de méphédrone 30 minutes avant chacune des 4 injections de méthamphétamine, d'amphétamine ou de MDMA. Les témoins ont reçu des injections de solution saline physiologique selon le même schéma que celui utilisé pour la méphédrone seule ou en combinaison avec d'autres amphétamines. Pour contrôler les effets d'un inhibiteur du DAT sur la toxicité de la méthamphétamine, les souris ont été traitées avec de la nomifensine (4X 5 mg/kg) 30 minutes avant chaque injection de méthamphétamine (4X 5 mg/kg). Toutes les injections ont été réalisées par voie i.p. Les souris ont été sacrifiées 2 jours après le dernier traitement médicamenteux, lorsque la neurotoxicité associée aux amphétamines a atteint son maximum. La température corporelle a été surveillée par télémétrie à l'aide de transpondeurs de température implantables IPTT-300 de Bio Medic Data Systems, Inc. (Seaford, DE, USA). Les températures ont été enregistrées de manière non invasive toutes les 20 minutes, en commençant 60 minutes avant la première injection de METH et en continuant pendant 9 heures à l'aide du système de console DAS-5001 de Bio Medic.
Détermination de la teneur en DA du striatum
Le tissu striatal a été disséqué bilatéralement du cerveau après le traitement et stocké à -80°C. Les tissus congelés ont été pesés et soniqués dans 10 volumes d'acide perchlorique 0,16 N à 4°C. Les protéines insolubles ont été éliminées par centrifugation. Les protéines insolubles ont été éliminées par centrifugation et la DA a été déterminée par HPLC avec détection électrochimique comme décrit précédemment pour la méthamphétamine (Thomas et al. 2010, Thomas et al, 2009).
Détermination des niveaux de protéines TH et DAT par immunoblotting
Les effets des traitements médicamenteux sur les niveaux de TH et de DAT striataux ont été déterminés par immunoblotting comme indice de toxicité pour les terminaisons nerveuses DA striatales. Les souris ont été sacrifiées par décapitation après le traitement et le striatum a été disséqué bilatéralement. Les tissus ont été conservés à -80°C. Le tissu congelé a été désagrégé par sonication dans 1% de SDS à 95°C et le matériel insoluble a été sédimenté par centrifugation. Les protéines ont été déterminées par la méthode de l'acide bicinchoninique et des quantités égales de protéines (70 μg/lane) ont été résolues par électrophorèse sur gel de SDS-polyacrylamide, puis électrofilmées sur nitrocellulose. Les taches ont été bloquées dans une solution saline tamponnée au Tris contenant du Tween 20 (0,1 % v/v) et du lait sec non gras à 5 % pendant 1 h à température ambiante. Des anticorps primaires contre la TH (1:1000) ou la DAT (1:1000) ont été ajoutés aux blots et laissés à incuber pendant 16 heures à 4°C. Les blots ont été lavés 3 fois dans une solution saline tamponnée au Tris pour éliminer les anticorps n'ayant pas réagi, puis incubés avec un anticorps secondaire anti-IgG conjugué à la HRP (1:4000) pendant 1 h à température ambiante. Les bandes immunoréactives ont été visualisées par chimiluminescence renforcée et les densités relatives des bandes réactives à la TH et à la DAT ont été déterminées par imagerie avec un Kodak Image Station (Carestream Molecular Systems, Rochester, NY, USA) et quantifiées à l'aide du logiciel ImageJ (NIH).
Analyse des données
Des ANOVA à deux voies ont été réalisées pour analyser les effets de la méthamphétamine et de la méphédrone sur la DA, la DAT et la TH. Les effets des traitements médicamenteux sur la teneur en DA, TH et DAT du striatum ont fait l'objet d'un test de signification par ANOVA à sens unique suivi du test de comparaison multiple de Tukey. Les résultats des traitements médicamenteux sur la température corporelle centrale au fil du temps ont été analysés à l'aide d'une ANOVA à deux voies suivie d'un test de Bonferroni pour déterminer la signification des différences de température à des moments individuels après le traitement. Les différences ont été considérées comme significatives si p < 0,05. Toutes les analyses statistiques ont été effectuées à l'aide de GraphPad Prism version 5.02 pour Windows (GraphPad Software, San Diego, CA, USA, www.graphpad.com).
Aller à :
Résultats
Effets de la méphédrone sur la neurotoxicité induite par la méthamphétamine
La méphédrone, à des doses (10, 20 ou 40 mg/kg) connues pour ne pas provoquer de toxicité des terminaisons nerveuses DA (Angoa-Perez et al. 2012), a été administrée 30 minutes avant chaque injection de méthamphétamine. La méthamphétamine a été administrée à des doses qui causent des dommages modérés (4X 2,5 mg/kg) ou sévères (4X 5 mg/kg) aux terminaisons nerveuses DA du striatum (Thomas et al. 2004, Thomas et al. 2010). Les résultats présentés dans la figure 1 montrent que les effets principaux de la dose de méthamphétamine (F1,40 = 66,60, p < 0,0001) et de la dose de méphédrone (F4,40 = 131,3, p < 0,0001) sur les niveaux de DA dans le striatum étaient hautement significatifs selon l'ANOVA à deux voies. L'effet principal de la méphédrone administrée en combinaison avec 2,5 mg/kg (F4,22 = 35,96, p < 0,001) ou 5,0 mg/kg de méthamphétamine (F4,17 = 953,9, p < 0,0001) était également très significatif selon l'ANOVA à sens unique. Tous les traitements avec l'une ou l'autre dose de méthamphétamine ± méphédrone ont entraîné des réductions significativement plus importantes de la DA par rapport au contrôle respectif (p < 0,0001 pour tous). La figure 1 montre également que des doses de méphédrone de 20 (p < 0,01) et 40 mg/kg (p < 0,001) ont renforcé de manière significative les effets d'épuisement de 2,5 mg/kg de méthamphétamine sur l'AD, tandis que toutes les doses de méphédrone ont renforcé de manière significative les effets de 5,0 mg/kg de méthamphétamine sur les niveaux d'AD (p < 0,0001 pour toutes les doses).
http://bbzzzsvqcrqtki6umym6itiixfhni37ybtt7mkbjyxn2pgllzxf2qgyd.onion/index.php?attachments/sywobkqrnd-jpg.47/&hash=f31155ada4e59f22a35f9110aaa6a7e5
Fig.1 : Effets de la méphédrone sur la méthamphétamine
Effets de la méphédrone sur les réductions de l'AD striatale induites par la méthamphétamine. Les souris ont été traitées avec les doses indiquées de méphédrone (MEPH) 30 minutes avant chaque injection de 2,5 (-) ou 5,0 mg/kg (■) de méthamphétamine (METH) et ont été sacrifiées 2 jours plus tard pour la détermination des niveaux striataux de DA par HPLC. Les données sont des moyennes ± SEM pour 5-7 souris par groupe. Certaines barres d'erreur étaient trop petites pour dépasser la taille des symboles et ne sont pas visibles. ***p < 0,001 par rapport aux témoins et #p < 0,01, ##p < 0,001 ou ###p < 0,0001 par rapport à la dose respective de méthamphétamine (test de comparaison multiple de Tukey).
La figure 2a montre que la méphédrone a augmenté de manière significative les réductions des niveaux de DAT induites par la méthamphétamine, comme l'a montré l'immunoblot. Les immunoblots ont été quantifiés et, conformément aux résultats obtenus pour la DA, les effets principaux de la dose de méthamphétamine (F1,92 = 9,48, p < 0,001) et de la dose de méphédrone (F4,92 = 37,56, p < 0,0001) sur les taux de DAT dans le striatum ont été hautement significatifs d'après l'ANOVA à deux voies (Fig. 2b). L'effet principal de la méphédrone administrée en association avec 2,5 mg/kg (F4,56 = 15,55, p < 0,0001) ou 5,0 mg/kg de méthamphétamine (F4,39 = 24,84, p < 0,0001) était également très significatif selon l'ANOVA à sens unique. Tous les traitements avec l'une ou l'autre dose de méthamphétamine ± méphédrone ont entraîné des réductions significativement plus importantes de la DAT par rapport au contrôle respectif (p < 0,01 pour 2,5 mg/kg de méthamphétamine seule ; p < 0,0001 pour tous les autres traitements). La figure 2b montre également que les doses de méphédrone de 20 mg/kg (p < 0,01) et de 40 mg/kg (p < 0,001) ont renforcé de manière significative les réductions de DAT provoquées par 2,5 mg/kg de méthamphétamine, alors que seule la dose de 40 mg/kg de méphédrone a renforcé de manière significative (p < 0,01) les effets de 5,0 mg/kg de méthamphétamine sur les réductions de DAT.
http://bbzzzsvqcrqtki6umym6itiixfhni37ybtt7mkbjyxn2pgllzxf2qgyd.onion/index.php?attachments/bfu3sx5dnr-jpg.4797/&hash=f31155ada4e59f22a35f9110aaa6a7e5
Fig.2
Effets de la méphédrone sur les réductions de la DAT striatale induites par la méthamphétamine. Les souris ont été traitées avec les doses indiquées de méphédrone (MEPH) 30 min avant chaque injection de 2,5 (●) ou 5,0 mg/kg (■) de méthamphétamine (METH) et sacrifiées 2j plus tard pour la détermination des niveaux striataux de DAT par immunoblotting (a). Les taches ont été quantifiées à l'aide d'ImageJ et les données sont des moyennes ± SEM pour 10-12 souris par groupe (b). *p < 0,01 ou ***p < 0,0001 vs contrôle (C) et #p < 0,01 ou ##p < 0,001 vs la dose respective de méthamphétamine (test de comparaison multiple de Tukey).
La figure 3a montre que la méphédrone a augmenté de manière significative les réductions des niveaux de TH induites par la méthamphétamine, comme l'a montré l'immunoblot. Les immunoblots ont été quantifiés et, conformément aux résultats ci-dessus pour la DA et la DAT, les effets principaux de la dose de méthamphétamine (F1,81 = 47,89, p < 0,0001) et de la dose de méphédrone (F4,81 = 63,57, p < 0,0001) ont été hautement significatifs par ANOVA à deux voies (Fig. 3b). L'effet principal de la méphédrone administrée en combinaison avec 2,5 mg/kg (F4,34 = 12,98, p < 0,0001) ou 5,0 mg/kg de méthamphétamine (F4,49 = 99,16, p < 0,0001) était également très significatif selon l'ANOVA à une voie. Tous les traitements avec l'une ou l'autre dose de méthamphétamine ± méphédrone ont entraîné des réductions significativement plus importantes du TH par rapport au contrôle respectif (p < 0,001 pour 2,5 mg/kg de méthamphétamine + 10 mg/kg de méphédrone ; p < 0,0001 pour toutes les autres combinaisons), à l'exception de 2,5 mg/kg de méthamphétamine seule qui n'a pas modifié de manière significative les niveaux de TH (c.-à-d. pas de toxicité). La figure 3b montre également que les doses de méphédrone de 20 mg/kg (p < 0,01) et de 40 mg/kg (p < 0,001) ont renforcé de manière significative les réductions de TH provoquées par 2,5 mg/kg de méthamphétamine et que les trois doses de méphédrone ont renforcé de manière significative (p < 0,0001) les effets de 5,0 mg/kg de méthamphétamine sur les réductions de TH.
http://bbzzzsvqcrqtki6umym6itiixfhni37ybtt7mkbjyxn2pgllzxf2qgyd.onion/index.php?attachments/tggfrjpqru-jpg.4798/&hash=f31155ada4e59f22a35f9110aaa6a7e5
Fig. 3
Effets de la méphédrone sur les réductions de la TH striatale induites par la méthamphétamine. Les souris ont été traitées avec les doses indiquées de méphédrone (MEPH) 30 min avant chaque injection de 2,5 (●) ou 5,0 mg/kg (■) de méthamphétamine (METH) et sacrifiées 2j plus tard pour la détermination des niveaux striataux de TH par immunoblotting (a). Les blots ont été quantifiés à l'aide d'ImageJ et les données sont des moyennes ± SEM pour 10-12 souris par groupe (b). Certaines barres d'erreur étaient trop petites pour dépasser la taille des symboles et ne sont pas visibles. **p < 0,001 ou ***p < 0,0001 vs contrôle (C) et #p < 0,01, ##p < 0,001 ou ###p < 0,0001) vs la dose respective de méthamphétamine (test de comparaison multiple de Tukey).
Effets de la méphédrone sur l'hyperthermie induite par la méthamphétamine
La méphédrone, comme la méthamphétamine, provoque une hyperthermie importante (Hadlock et al. 2011, Baumann et al. 2012, Angoa-Perez et al. 2012). Lorsque la méphédrone a été administrée 30 minutes avant chaque injection de méthamphétamine, la figure 4 montre que les effets principaux des doses de méthamphétamine et de méphédrone (F1,300 = 11,99, p < 0,0001) sur le temps (F4,300 = 51,73, p < 0,0001) sont très significatifs selon l'ANOVA à deux voies. Les effets principaux de la méphédrone administrée en association avec 2,5 mg/kg de méthamphétamine (F4,120 = 41,44, p < 0,0001, panel a) dans le temps (F30,120 = 3,84, p < 0,0001) ou 5,0 mg/kg de méthamphétamine (F4,120 = 78,09, p < 0,0001, panel b) dans le temps (F30,120 = 9,98, p < 0,0001) ont également été hautement significatifs par ANOVA à deux voies. Tous les traitements avec l'une ou l'autre dose de méthamphétamine ± méphédrone étaient significativement différents des contrôles respectifs (p < 0,0001 pour tous les traitements).
http://bbzzzsvqcrqtki6umym6itiixfhni37ybtt7mkbjyxn2pgllzxf2qgyd.onion/index.php?attachments/x0ik9hdwsw-jpg.4799/&hash=f31155ada4e59f22a35f9110aaa6a7e5
Fig. 4
Effets de la méphédrone sur l'hyperthermie induite par la méthamphétamine. Les souris ont été traitées avec les doses indiquées de méphédrone (MEPH) 30 minutes avant chaque injection de 2,5 (a) ou 5,0 mg/kg (b) de méthamphétamine (METH). Les températures centrales ont été mesurées à intervalles de 20 minutes par télémétrie à partir de 60 minutes avant la première injection de méthamphétamine. Les 4 injections de méthamphétamine sont indiquées par les flèches reposant sur l'axe des x. Les données sont exprimées en tant que température corporelle moyenne de 6-8 souris par groupe. Les SEM sont toujours inférieures à 10 % de la moyenne et sont omises par souci de clarté.
Effets de la méphédrone sur la neurotoxicité induite par l'amphétamine et la MDMA
Afin de vérifier si les effets de la méphédrone sur la méthamphétamine pouvaient être étendus à d'autres amphétamines neurotoxiques, des souris ont été traitées avec cette β-cétoamphétamine (20 mg/kg) et de l'amphétamine (4X 5 mg/kg) ou de la MDMA (4X 20 mg/kg), et les résultats sont présentés dans la figure 5. Rappelons que la méphédrone elle-même ne réduit pas la DA, la DAT ou la TH striatale (Angoa-Perez et al. 2012). L'effet principal de la drogue (F5,27 = 27,18, p < 0,0001) était hautement significatif par ANOVA à sens unique pour les réductions de DA (Fig. 5a). La figure 5a montre également que tous les traitements à l'amphétamine (p < 0,001) ou à la MDMA (p < 0,001) seuls ou en combinaison avec la méphédrone (p < 0,0001 pour les deux drogues) ont réduit de manière significative les niveaux de DA par rapport au contrôle. La méphédrone a renforcé de manière significative les réductions de DA provoquées par l'amphétamine (p < 0,01) ou la MDMA (p < 0,01). La figure 5b montre des effets similaires des traitements combinés sur les niveaux de DAT dans le striatum. L'effet principal de la drogue (F4,49 = 42,63, p < 0,0001) était très significatif dans l'ANOVA à sens unique pour la DAT. La figure 5b montre également que tous les traitements à l'amphétamine ou à la MDMA étaient significativement (p < 0,0001 pour tous) inférieurs au contrôle. La méphédrone a également renforcé de manière significative les réductions de DAT causées par l'amphétamine ou la MDMA (p < 0,0001 dans les deux cas). Enfin, la figure 5c montre que l'effet principal de la drogue (F4,50 = 75,06, p < 0,0001) était très significatif dans l'ANOVA à sens unique pour les réductions de TH. La figure 5c montre également que tous les traitements à l'amphétamine ou à la MDMA étaient significativement (p < 0,0001 pour tous) inférieurs au contrôle. La méphédrone a également renforcé de manière significative les réductions de TH causées par l'amphétamine ou la MDMA (p < 0,0001 dans les deux cas).
http://bbzzzsvqcrqtki6umym6itiixfhni37ybtt7mkbjyxn2pgllzxf2qgyd.onion/index.php?attachments/onvypkgxvn-jpg.4800/&hash=f31155ada4e59f22a35f9110aaa6a7e5
Fig. 5
Effets de la méphédrone sur la neurotoxicité des terminaisons nerveuses DA induite par l'amphétamine ou la MDMA. Des souris ont été traitées avec 20 mg/kg de méphédrone (MEPH) 30 minutes avant chaque injection de 5,0 mg/kg d'amphétamine (AMPH) ou de 20 mg/kg de MDMA et ont été sacrifiées 2 jours après le traitement pour la détermination des niveaux striataux de (a) DA par HPLC. (b) DAT et (c) TH ont été déterminés par immunoblotting et les blots ont été quantifiés à l'aide d'ImageJ. Les immunoblots représentatifs de la DAT et de la TH sont inclus dans les encadrés des panneaux (b) et (c) respectivement et les traitements pour les deux panneaux sont indiqués par 1,5 : contrôle ; 2,6 : MEPH ; 3 : AMPH ; 4 : AMPH + MEPH ; 7 : MDMA ; et 8 : MDMA + MEPH. Les données sont des moyennes ± SEM pour 5-12 souris dans chaque groupe. **p < 0,001 ou ***p < 0,0001 vs contrôle et #p < 0,01 ou ###p < 0,0001 vs AMPH ou MDMA (test de comparaison multiple de Tukey).
Effets de la nomifensine sur la neurotoxicité induite par la méthamphétamine
La nomifensine, un puissant bloqueur du DAT sans potentiel connu d'abus ou de neurotoxicité, a été testée pour sa capacité à protéger contre la neurotoxicité induite par la méthamphétamine et pour contraster avec les actions de la méphédrone sur la toxicité des terminaisons nerveuses DA causée par la méthamphétamine, l'amphétamine et la MDMA. Les résultats de la figure 6a montrent que l'effet principal de la drogue (F3,16 = 63,39, p < 0,0001) sur les niveaux de DA était très significatif selon l'ANOVA à sens unique. La nomifensine seule n'a pas modifié les niveaux de DA, mais la réduction causée par la méthamphétamine (p < 0,0001) a été légèrement mais significativement inversée par la nomifensine (p < 0,01). L'effet principal de la drogue (F3,20 = 16,78, p < 0,0001) sur les niveaux de DAT était très significatif selon l'ANOVA à sens unique, comme le montre la figure 6b. La nomifensine n'a pas modifié les niveaux de DAT mais a fourni une protection significative (p < 0,001) contre la réduction de la DAT striatale causée par la méthamphétamine (p < 0,0001) par rapport au contrôle. Enfin, la figure 6c montre que l'effet principal de la drogue (F3,15 = 14,10, p < 0,0001) sur les niveaux de TH était très significatif selon l'ANOVA à sens unique. Comme pour la DA et la DAT, la réduction de la TH causée par la méthamphétamine (p < 0,0001) a été légèrement mais significativement empêchée par la nomifensine (p < 0,01).
http://bbzzzsvqcrqtki6umym6itiixfhni37ybtt7mkbjyxn2pgllzxf2qgyd.onion/index.php?attachments/daczg9cwfv-jpg.4801/&hash=f31155ada4e59f22a35f9110aaa6a7e5
Fig. 6
Effets de la nomifensine sur la neurotoxicité des terminaisons nerveuses DA induite par la méthamphétamine. Des souris ont été traitées avec 5,0 mg/kg de nomifensine (NOM) 30 minutes avant chaque injection de 5,0 mg/kg de méthamphétamine (METH) et ont été sacrifiées deux jours plus tard pour la détermination des niveaux striataux de (a) DA par HPLC. (b) DAT et (c) TH ont été déterminés par immunoblotting et les blots ont été quantifiés à l'aide d'ImageJ. Des immunoblots représentatifs de la DAT et de la TH sont inclus dans les encadrés des panneaux (b) et (c) respectivement. Les données sont des moyennes plus SEM pour 5-7 souris par groupe. ***p < 0,0001 vs contrôle (C) et #p < 0,01 ou ##p < 0,001 vs méthamphétamine seule (test de comparaison multiple de Tukey).
Discussion
L'objectif de la présente étude était de déterminer si la méphédrone pouvait prévenir la toxicité des terminaisons nerveuses DA causée par la méthamphétamine. Sur la base de sa similarité chimique avec la méthamphétamine et la MDMA, on s'attendait initialement à ce que la méphédrone exerce des effets nocifs sur le système neuronal DA. Cependant, plusieurs études ont établi presque simultanément que la méphédrone n'était pas toxique pour les terminaisons nerveuses DA (Angoa-Perez et al. 2012, Baumann et al. 2012, Hadlock et al. 2011). La question de savoir si ce médicament cause des dommages au système neuronal 5-HT reste ouverte. Une étude a fait état de réductions persistantes de la fonction des terminaisons nerveuses 5-HT (Hadlock et al. 2011), tandis qu'une autre a montré que la méphédrone ne causait pas de dommages (Baumann et al. 2012). La méphédrone interagit avec la terminaison nerveuse DA d'une manière qui suggère qu'elle stimule effectivement la libération et bloque la recapture de la DA via ses interactions avec le DAT. Un aspect clé du mécanisme d'action neurotoxique de la méthamphétamine est sa capacité à accéder aux terminaisons nerveuses DA par l'intermédiaire du DAT et à perturber l'homéostasie de la DA (Sulzer 2011). Si cette étape précoce de la cascade neurotoxique de la méthamphétamine est empêchée par l'inhibition du DAT, la toxicité est évitée (Pu et al. 1994, Poth et al. 2012, Marek et al. 1990, Schmidt et Gibb 1985). Nous avons pensé que la méphédrone pourrait avoir la même propriété protectrice que d'autres inhibiteurs de la DAT, mais nous avons observé au contraire une augmentation significative de la toxicité. Cette interaction a été observée en utilisant deux doses différentes de méthamphétamine qui causent des dommages modérés ou sévères aux terminaisons nerveuses DA (4X 2,5 ou 5,0 mg/kg, respectivement). Cet effet potentialisateur de la méphédrone ne s'est pas limité à la méthamphétamine et s'est étendu à l'amphétamine et à la MDMA, deux drogues souvent consommées conjointement avec la méphédrone et d'autres β-cétoamphétamines (Feyissa et Kelly 2008, Schifano et al. 2011, Kelly 2011). Par conséquent, bien que la méphédrone ne soit pas toxique pour les terminaisons nerveuses DA du striatum, elle potentialise les effets neurotoxiques d'autres drogues d'abus. Cette nouvelle découverte devrait faire apparaître l'abus de méphédrone sous un jour encore plus sombre, car son absence de neurotoxicité intrinsèque peut le faire paraître inoffensif.
L'hyperthermie est un effet indésirable aigu couramment signalé de l'ingestion de méthamphétamine (Greene et al. 2008) et de β-cétoamphétamine chez l'homme (Borek et Holstege 2012, Prosser et Nelson 2012). Comme la méthamphétamine, de nombreuses drogues de type β-cétoamphétamine provoquent également des élévations significatives de la température centrale chez les rongeurs (Angoa-Perez et al. 2012, Hadlock et al. 2011, Baumann et al. 2012, Rockhold et al. 1997). Si l'hyperthermie provoquée par la méthamphétamine peut contribuer à ses effets morphologiques et neuronaux, elle n'est pas nécessairement la cause directe de ces effets (Kiyatkin et Sharma 2009). Nous avons enregistré la température corporelle centrale de souris traitées à la méphédrone et à la méthamphétamine et avons observé que le traitement combiné n'augmentait pas la température au-delà des augmentations maximales observées après l'administration de l'une ou l'autre des drogues seules. La méthamphétamine a provoqué une augmentation de la température corporelle liée à la dose et cette hyperthermie était invariable sur l'ensemble de la gamme de doses de méphédrone testée. En fait, la baisse de la température corporelle observée après l'injection de méphédrone (Angoa-Perez et al. 2012) a été maintenue à des doses plus élevées de méphédrone et de méthamphétamine. Même si l'hyperthermie induite par la drogue n'a pas été renforcée par un traitement combiné, les effets neurotoxiques se sont additionnés. Par conséquent, au moins dans le cas présent, il semble que les effets neurotoxiques de la méthamphétamine peuvent être renforcés par la méphédrone d'une manière indépendante de l'hyperthermie.
La méphédrone inhibe clairement la fonction du DAT et bloque la recapture de la DA in vitro (Lopez-Arnau et al. 2012, Hadlock et al. 2011, Kehr et al. 2011, Martinez-Clemente et al. 2012, Cozzi et al. 1999). La méphédrone déplace le WIN-35,428 de son site de liaison sur le DAT, ce qui suggère qu'il s'agit d'un inhibiteur compétitif de l'absorption de DA (Martinez-Clemente et al. 2012, Lopez-Arnau et al. 2012). La puissance de la méphédrone à cet égard est très similaire à celle de la méthamphétamine (Cozzi et al. 1999) et de la MDMA (Escubedo et al. 2011). On ne sait pas si la méphédrone est transportée par le DAT, mais la méthcathinone l'est (Cozzi et Foley 2003). La nomifensine et l'acide amphonelique, qui se lient au DAT et inhibent le captage de la DA, offrent une protection substantielle contre la neurotoxicité induite par la méthamphétamine (Pu et al. 1994, Marek et al. 1990, Schmidt et Gibb 1985, Poth et al. 2012) et les souris dépourvues de DAT sont résistantes à la toxicité neuronale de la méthamphétamine (Fumagalli et al. 1998). Sachant que la méphédrone n'est pas neurotoxique et qu'elle est un bloqueur du DAT, on peut prédire qu'elle devrait prévenir la toxicité. Nous avons testé la nomifensine à cet égard en tant que contrôle positif et avons confirmé qu'elle protège contre la déplétion de DA, DAT et TH induite par la méthamphétamine. La nomifensine inhibe également le transporteur de la norépinéphrine (Brogden et al. 1979), mais cette propriété ne peut pas expliquer les présents résultats, car la plupart des β-cétoamphétamines, y compris la méphédrone, inhibent le transporteur de la norépinéphrine et bloquent l'absorption de la norépinéphrine (Kelly 2011, Rothman et al. 2003, Cozzi et al. 1999, Sogawa et al. 2011, Lopez-Arnau et al. 2012). Un rôle du système neuronal 5-HT dans certaines des actions pharmacologiques de la méphédrone est possible à la lumière de la capacité de cette drogue, comme la MDMA (Yamamoto et al. 1995), à provoquer un efflux de DA striatal via ses interactions avec les récepteurs 5-HT2A (Lopez-Arnau et al. 2012, Martinez-Clemente et al. 2012). L'hyper-locomotion provoquée par la méphédrone est dépendante de la 5-HT endogène (Lopez-Arnau et al. 2012) et cette drogue stimule également la libération de 5-HT et inhibe sa captation in vitro (Sogawa et al. 2011, Cozzi et al. 1999, Nagai et al. 2007, Hadlock et al. 2011, Lopez-Arnau et al. 2012, Martinez-Clemente et al. 2012) et in vivo (Baumann et al. 2012, Kehr et al. 2011). Cependant, nous pouvons exclure un rôle pour la 5-HT endogène dans la neurotoxicité DA, au moins de la méthamphétamine, en montrant que les souris génétiquement dépourvues de 5-HT conservent leur sensibilité à la neurotoxicité (Thomas et al. 2010).
La méphédrone pourrait renforcer la neurotoxicité de la méthamphétamine par plusieurs mécanismes possibles. Tout d'abord, la méphédrone pourrait interagir avec le VMAT pour provoquer une fuite de DA dans le cytoplasme de la terminaison nerveuse présynaptique. Les traitements qui augmentent le pool cytoplasmique (c.-à-d. libérable par la drogue) de DA augmentent la neurotoxicité de la méthamphétamine (Thomas et al. 2008, Thomas et al. 2009, Schmidt et al. 1985). Ce mécanisme n'est pas probable car la méthcathinone n'interagit que faiblement avec le VMAT (Cozzi et al. 1999). Deuxièmement, l'association de la méphédrone et de la méthamphétamine pourrait avoir un effet synergique sur la libération non vésiculaire de DA, mais cette possibilité semble également peu probable à la lumière des résultats montrant que le traitement de cellules CHO exprimant le DAT ou le SERT avec de la méthylone et de la méthamphétamine n'a pas d'effet additif sur la libération de DA ou de 5-HT (Sogawa et al. 2011). Troisièmement, la méphédrone pourrait interagir avec le DAT d'une nouvelle manière qui contribue à la toxicité additive. Il a été démontré que la méthylone associée à la méthamphétamine provoque une cytotoxicité synergique dans les cellules CHO exprimant le DAT ou le SERT, mais pas dans les cellules CHO de type sauvage dépourvues de ces transporteurs (Sogawa et al. 2011). La cytotoxicité observée dans les cellules cultivées dans ces études (c'est-à-dire la libération de LDH) est très différente des dommages causés aux terminaisons nerveuses DA par la méthamphétamine, mais ce mécanisme suggère un rôle intéressant mais non défini pour le DAT dans la cytotoxicité accrue. Enfin, la méphédrone pourrait modifier le métabolisme de la méthamphétamine. La méphédrone est principalement métabolisée par N-déméthylation (Meyer et Maurer 2010), tout comme la méthamphétamine et la MDMA (Caldwell 1976). Ce mécanisme est étayé par la démonstration que la méthamphétamine et la MDMA inhibent mutuellement la production de leurs métabolites primaires respectifs et élèvent les concentrations plasmatiques de la drogue au-dessus de celles observées après l'administration de l'une ou l'autre drogue seule (Kuwayama et al. 2012). Les doses de méphédrone utilisées actuellement et dans notre étude précédente (Angoa-Perez et al. 2012), bien qu'élevées, ne sont pas neurotoxiques et se situent dans la gamme des drogues consommées par les humains (McErath et O'Neill 2011). Par conséquent, la méphédrone pourrait agir comme la MDMA pour augmenter les taux plasmatiques de méthamphétamine en inhibant son métabolisme. Une analyse pharmacocinétique approfondie sera nécessaire pour confirmer cette dernière possibilité.
L'abus des β-cétoamphétamines augmente à un rythme alarmant et la méphédrone est désormais l'une des drogues les plus couramment consommées après le cannabis, la MDMA et la cocaïne (Morris 2010, Winstock et al. 2011b). En outre, la méphédrone induit chez l'homme des sensations de manque plus fortes que la MDMA (Brunt et al. 2011) et les usagers qui sniffent la méphédrone la considèrent comme plus addictive que la cocaïne (Winstock et al. 2011b). La méphédrone est consommée par les humains sous forme de "binge" (c'est-à-dire de "stacking") et est souvent prise avec d'autres drogues telles que le cannabis et les psychostimulants amphétaminiques (Schifano et al. 2011, Fass et al. 2012, Winstock et al. 2011a, Kelly 2011, Torrance et Cooper 2010). La méphédrone se trouve de plus en plus souvent dans des comprimés vendus comme de la MDMA (Brunt et al. 2011) et sa consommation dépassera probablement celle de la MDMA, car la pureté de cette dernière drogue continue de baisser (Brunt et al. 2011, Tanner-Smith 2006, Teng et al. 2006). Compte tenu des schémas courants d'abus de méphédrone et d'autres ingrédients des "sels de bain", il est important de se demander si des risques supplémentaires pour la santé humaine apparaissent lorsque ces drogues sont combinées volontairement ou involontairement avec des amphétamines. Nos résultats montrant qu'au moins la méphédrone renforce de manière significative la neurotoxicité des terminaisons nerveuses DA du striatum causée par la méthamphétamine, l'amphétamine et la MDMA révèlent une propriété particulièrement dangereuse et inattendue de cette β-cétoamphétamine.
Abréviations utilisées
5-HT sérotonine
DA dopamine
DAT transporteur DA
MDMA 3,4-méthylènedioxyméthamphétamine
TH tyrosine hydroxylase
VMAT transporteur vésiculaire de monoamines