A mefedrona (4-metilmetcatinona) é uma droga de abuso estimulante β-cetoanfetamina com estreitas semelhanças estruturais e mecânicas com a metanfetamina. Uma das acções mais potentes associadas à mefedrona é a capacidade de estimular a libertação de dopamina (DA) e bloquear a sua recaptação através da sua interação com o transportador de dopamina (DAT). Embora a mefedrona não provoque toxicidade nas terminações nervosas DA, a sua capacidade de atuar como bloqueador do DAT poderia proporcionar proteção contra a neurotoxicidade induzida pela metanfetamina, tal como outros inibidores do DAT. Para testar esta possibilidade, os ratinhos foram tratados com mefedrona (10, 20 ou 40 mg/kg) antes de cada injeção de um regime neurotóxico de metanfetamina (4 injecções de 2,5 ou 5,0 mg/kg com intervalos de 2 horas). A integridade das terminações nervosas DA do striatum foi avaliada através de medidas dos níveis de DA, DAT e tirosina hidroxilase. A toxicidade moderada a grave da DA associada às diferentes doses de metanfetamina não foi evitada por nenhuma dose de mefedrona, mas foi, de facto, significativamente aumentada. A hipertermia causada pelo tratamento combinado com mefedrona e metanfetamina foi a mesma que a observada com qualquer um dos fármacos isoladamente. A mefedrona também aumentou os efeitos neurotóxicos da anfetamina e da MDMA nas terminações nervosas DA. Em contrapartida, a nomifensina protegeu contra a neurotoxicidade induzida pela metanfetamina. Uma vez que a mefedrona aumenta a neurotoxicidade da metanfetamina, os presentes resultados sugerem que interage com o DAT de uma forma diferente da de outros inibidores típicos do DAT. Os efeitos relativamente inócuos da mefedrona, por si só, nas terminações nervosas da DA ocultam uma interação potencialmente perigosa com drogas que são frequentemente utilizadas em conjunto com ela, levando a uma maior neurotoxicidade.
A mefedrona (4-metilmetcatinona) é um derivado da catinona e análogo estrutural da metanfetamina e da 3,4-metilenodioxi-metanfetamina (MDMA). A mefedrona é um dos ingredientes psicoactivos dos "sais de banho", juntamente com outros compostos, como a metilona, a butilona e a 3,4-metilenodioxipirovalerona (MDPV). As β-cetoanfetaminas estão a ser consumidas a taxas crescentes devido, em grande parte, à disponibilidade altamente restrita dos precursores necessários para a síntese de metanfetamina e MDMA em laboratórios clandestinos e à correspondente redução da sua pureza (Winstock et al. 2011b, Brunt et al. 2011). À medida que o abuso de β-cetoanfetaminas continua a aumentar, a lista dos seus efeitos adversos cresceu para incluir complicações cardiovasculares, agitação, insónia, psicose e depressão (Schifano et al. 2011, Prosser e Nelson 2012).
Como congéneres químicos da metanfetamina e da MDMA, não é surpreendente que as β-cetoanfetaminas tenham muitos dos mesmos efeitos que estas drogas anteriores no sistema nervoso central. Por exemplo, estas drogas bloqueiam os transportadores de dopamina (DA) e de serotonina (5-HT) (DAT e SERT, respetivamente) (Cozzi et al. 1999, Rothman et al. 2003, Fleckenstein et al. 2000, Lopez-Arnau et al. 2012) e estimulam a libertação de monoaminas in vitro (Kalix e Glennon 1986, Gygi et al. 1997, Rothman et al. 2003) e in vivo (Gygi et al. 1997, Kehr et al. 2011). A metcatinona provoca reduções persistentes da atividade da triptofano hidroxilase e da tirosina hidroxilase (TH) e depleção de DA e 5-HT (Gygi et al. 1997, Gygi et al. 1996, Sparago et al. 1996). Estudos de imagem PET em consumidores abstinentes de metcatinona revelaram uma redução da densidade do DAT no estriado, o que sugere uma perda de terminais DA (McCann et al. 1998). A estimulação simultânea da libertação de DA e a inibição da sua captação espelham os elementos críticos subjacentes à neurotoxicidade associada à metanfetamina (Kuhn et al. 2008, Yamamoto e Bankson 2005, Cadet et al. 2007, Fleckenstein et al. 2007).
Nós (Angoa-Perez et al. 2012) e outros (Baumann et al. 2012, Hadlock et al. 2011) investigámos recentemente a possibilidade de a mefedrona poder causar neurotoxicidade como a metanfetamina e a MDMA. Surpreendentemente, a mefedrona não era tóxica para as terminações nervosas DA do estriado (Hadlock et al. 2011, Baumann et al. 2012, Angoa-Perez et al. 2012). A questão de saber se a mefedrona danifica as terminações nervosas 5-HT continua por resolver, uma vez que um estudo documentou efeitos positivos (Hadlock et al. 2011), enquanto outro foi negativo (Baumann et al. 2012). À luz do efeito relativamente benigno da mefedrona nas terminações nervosas DA e considerando as suas propriedades como bloqueador DAT, colocámos a hipótese de que poderia realmente proteger o sistema neuronal DA dos efeitos neurotóxicos da metanfetamina, tal como se sabe que ocorre com outros bloqueadores DAT, como o ácido anfonélico (Pu et al. 1994, Schmidt e Gibb 1985, Marek et al. 1990) e nomifensina (Poth et al. 2012). Atualmente, verificamos que a mefedrona aumenta significativamente a neurotoxicidade da metanfetamina. Este efeito estende-se à anfetamina e à MDMA, drogas que são frequentemente consumidas em conjunto com a mefedrona (Feyissa e Kelly 2008, Schifano et al. 2011). Estes resultados surpreendentes lançam uma nova luz sobre o abuso da mefedrona e tornam urgente o reconhecimento desta propriedade subtil e perigosa desta β-cetoanfetamina.
Materiais e métodos
Drogas e reagentes
O cloridrato de mefedrona e a 3,4-metilenodioximetanfetamina (MDMA) foram obtidos no Programa de Fornecimento de Drogas do NIDA Research Resources. (+) O cloridrato de metanfetamina, o maleato de nomifensina, o sulfato de d-anfetamina, o pentobarbital, a DA e todos os tampões e reagentes de HPLC foram adquiridos à Sigma-Aldrich (St. Louis, MO, EUA). Os kits de ensaio de proteínas de ácido bicinconínico foram obtidos da Pierce (Rockford, IL, EUA). Os anticorpos policlonais contra a TH do rato foram produzidos conforme descrito anteriormente (Kuhn e Billingsley 1987). Os anticorpos monoclonais contra a DAT do rato foram generosamente fornecidos pela Dra. Roxanne A. Vaughan (Universidade do Dakota do Norte, Grand Forks, ND, EUA). Os anticorpos secundários anti-IgG conjugados com HRP foram fornecidos por Jackson ImmunoResearch Laboratories, Inc. (West Grove, PA, EUA).
Animais
As fêmeas de ratinhos C57BL/6 (Harlan, Indianapolis, IN, EUA), com um peso de 20-25 g na altura da experiência, foram alojadas 5 por gaiola em gaiolas grandes com caixa de sapatos, numa sala com luz (12 h de luz/obscuridade) e temperatura controlada. Foram utilizadas ratinhas fêmeas porque se sabe que são muito sensíveis aos danos neuronais provocados pelas anfetaminas neurotóxicas e para manter a coerência com os nossos estudos anteriores sobre a neurotoxicidade da metanfetamina (Thomas et al. 2010, Thomas et al. 2008, Thomas et al. 2009). Os ratinhos tinham livre acesso a alimentos e água. O Comité Institucional de Cuidados e Utilização da Wayne State University aprovou os cuidados com os animais e os procedimentos experimentais. Todos os procedimentos também estavam em conformidade com o Guia do NIH para o Cuidado e Uso de Animais de Laboratório.
Procedimentos farmacológicos, fisiológicos e comportamentais
Os ratinhos foram tratados com mefedrona utilizando um regime de tratamento em "binge" composto por 4 injecções de 10, 20 ou 40 mg/kg com um intervalo de 2 horas entre cada injeção. Este regime de tratamento em "binge", quando utilizado para injetar anfetaminas substituídas e derivados da catinona, provoca lesões extensas nas terminações nervosas DA. As doses de mefedrona atualmente utilizadas demonstraram anteriormente não serem tóxicas para as terminações nervosas DA (Angoa-Perez et al. 2012). Os ratinhos foram tratados com metanfetamina (4X 2,5 ou 5 mg/kg), anfetamina (4X 5 mg/kg) ou MDMA (4X 20 mg/kg) isoladamente ou em combinação com mefedrona. Quando tratados com duas drogas, os ratos receberam uma injeção de mefedrona 30 minutos antes de cada uma das 4 injecções de metanfetamina, anfetamina ou MDMA. Os controlos receberam injecções de soro fisiológico no mesmo horário utilizado para a mefedrona isolada ou em combinação com outras anfetaminas. Como controlo dos efeitos de um inibidor do DAT na toxicidade da metanfetamina, os ratinhos foram tratados com nomifensina (4X 5 mg/kg) 30 minutos antes de cada injeção de metanfetamina (4X 5 mg/kg). Todas as injecções foram administradas por via i.p.. Os ratinhos foram sacrificados 2 dias após o último tratamento medicamentoso, quando a neurotoxicidade associada à anfetamina atingiu o seu máximo. A temperatura corporal foi monitorizada por telemetria utilizando transponders de temperatura implantáveis IPTT-300 da Bio Medic Data Systems, Inc. (Seaford, DE, EUA). As temperaturas foram registadas de forma não invasiva a cada 20 minutos, começando 60 minutos antes da primeira injeção de METH e continuando durante as 9 horas seguintes, utilizando o sistema de consola DAS-5001 da Bio Medic.
Determinação do teor de DA no estriado
O tecido estriado foi dissecado bilateralmente do cérebro após o tratamento e armazenado a -80°C. Os tecidos congelados foram pesados e submetidos a sonicação em 10 volumes de ácido perclórico 0,16 N a 4°C. A proteína insolúvel foi removida por centrifugação e a DA foi determinada por HPLC com deteção eletroquímica, tal como descrito anteriormente para a metanfetamina (Thomas et al. 2010, Thomas et al, 2009).
Determinação dos níveis de proteínas TH e DAT por immunoblotting
Os efeitos dos tratamentos medicamentosos nos níveis estriatais de TH e DAT foram determinados por immunoblotting como um índice de toxicidade para as terminações nervosas DA estriatais. Os ratos foram sacrificados por decapitação após o tratamento e o estriado foi dissecado bilateralmente. O tecido foi armazenado a -80°C. O tecido congelado foi desfeito por sonicação em SDS a 1% a 95°C e o material insolúvel foi sedimentado por centrifugação. A proteína foi determinada pelo método do ácido bicinchonínico e quantidades iguais de proteína (70 μg/lano) foram resolvidas por eletroforese em gel de SDS-poliacrilamida e depois electroblotadas para nitrocelulose. Os blots foram bloqueados em solução salina tamponada com Tris contendo Tween 20 (0,1% v/v) e 5% de leite seco magro durante 1 h à temperatura ambiente. Os anticorpos primários contra TH (1:1000) ou DAT (1:1000) foram adicionados aos blots e deixados a incubar durante 16 horas a 4°C. Os blots foram lavados 3 vezes em solução salina tamponada com Tris para remover os anticorpos que não reagiram e depois incubados com anticorpo secundário anti-IgG conjugado com HRP (1:4000) durante 1 h à temperatura ambiente. As bandas imunorreactivas foram visualizadas por quimioluminescência reforçada e as densidades relativas das bandas reactivas a TH e DAT foram determinadas por imagem com uma Kodak Image Station (Carestream Molecular Systems, Rochester, NY, EUA) e quantificadas utilizando o software ImageJ (NIH).
Análise dos dados
Foram efectuadas ANOVAs de duas vias para analisar os efeitos da dose de metanfetamina versus mefedrona na DA, DAT e TH. Os efeitos dos tratamentos com drogas no conteúdo estriatal de DA, TH e DAT foram testados quanto à significância por ANOVA de uma via seguida do teste de comparação múltipla de Tukey. Os resultados dos tratamentos medicamentosos na temperatura corporal central ao longo do tempo foram analisados usando uma ANOVA de duas vias seguida pelo teste de Bonferroni para determinar a significância das diferenças de temperatura em momentos individuais após o tratamento. As diferenças foram consideradas significativas se p < 0,05. Todas as análises estatísticas foram realizadas usando o GraphPad Prism versão 5.02 para Windows (GraphPad Software, San Diego, CA, EUA, www.graphpad.com).
Ir para:
Resultados
Efeitos da mefedrona na neurotoxicidade induzida pela metanfetamina
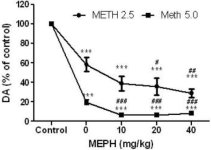
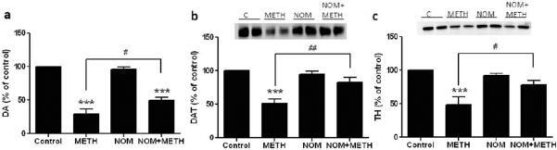
A mefedrona, em doses (10, 20 ou 40 mg/kg) que se sabe não causarem toxicidade na terminação nervosa DA (Angoa-Perez et al. 2012) foi administrada 30 minutos antes de cada injeção de metanfetamina. A metanfetamina foi administrada em doses que causam danos moderados (4X 2,5 mg/kg) ou graves (4X 5 mg/kg) nas terminações nervosas DA do estriado (Thomas et al. 2004, Thomas et al. 2010). Os resultados apresentados na Fig. 1 mostram que os principais efeitos da dose de metanfetamina (F1,40 = 66.60, p <0.0001) e da dose de mefedrona (F4,40 = 131.3, p <0.0001) nos níveis de DA no estriado foram altamente significativos pela ANOVA de duas vias. O efeito principal da mefedrona administrada em combinação com 2.5 mg / kg (F4,22 = 35.96, p <0.001) ou 5.0 mg / kg de metanfetamina (F4,17 = 953.9, p <0.0001) também foi altamente significativo por ANOVA unidirecional. Todos os tratamentos com qualquer dose de metanfetamina ± mefedrona causaram reduções significativamente maiores em DA em comparação com o respetivo controle (p <0.0001 para todos). A Fig. 1 mostra também que as doses de mefedrona de 20 (p < 0,01) e 40 mg/kg (p < 0,001) aumentaram significativamente os efeitos depletores de 2,5 mg/kg de metanfetamina na AD, enquanto todas as doses de mefedrona aumentaram significativamente os efeitos de 5,0 mg/kg de metanfetamina nos níveis de AD (p < 0,0001 para todos).
Fig.1
Efeitos da mefedrona nas reduções da DA estriatal induzidas pela metanfetamina. Os ratos foram tratados com as doses indicadas de mefedrona (MEPH) 30 min antes de cada injeção de 2.5 (-) ou 5.0 mg / kg (■) de metanfetamina (METH) e sacrificados 2d mais tarde para determinação dos níveis estriatais de DA por HPLC. Os dados são médios ± SEM para 5-7 ratos por grupo. Algumas barras de erro eram demasiado pequenas para exceder o tamanho dos símbolos e não aparecem visíveis. ***p < 0,001 vs controlos e #p < 0,01, ##p < 0,001 ou ###p < 0,0001 vs a respectiva dose de metanfetamina (teste de comparação múltipla de Tukey).
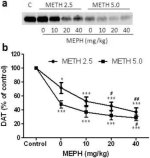
Fig. 2a mostra que a mefedrona aumentou significativamente as reduções induzidas pela metanfetamina nos níveis de DAT, conforme determinado por immunoblotting. Os immunoblots foram quantificados e, de acordo com os resultados para DA, os principais efeitos da dose de metanfetamina (F1,92 = 9.48, p <0.001) e da dose de mefedrona (F4,92 = 37.56, p <0.0001) nos níveis de DAT no estriado foram altamente significativos por ANOVA de duas vias (Fig. 2b). O efeito principal da mefedrona administrada em combinação com 2.5 mg / kg (F4,56 = 15.55, p <0.0001) ou 5.0 mg / kg de metanfetamina (F4,39 = 24.84, p <0.0001) também foi altamente significativo por ANOVA unidirecional. Todos os tratamentos com qualquer dose de metanfetamina ± mefedrona causaram reduções significativamente maiores no DAT em comparação com o respetivo controle (p <0.01 para 2.5 mg / kg de metanfetamina sozinho; p <0.0001 para todos os outros tratamentos). A Fig. 2b mostra também que as doses de mefedrona de 20 mg/kg (p < 0,01) e 40 mg/kg (p < 0,001) aumentaram significativamente as reduções do DAT causadas por 2,5 mg/kg de metanfetamina, ao passo que apenas a dose de 40 mg/kg de mefedrona aumentou significativamente (p < 0,01) os efeitos de 5,0 mg/kg de metanfetamina nas reduções do DAT.
Fig.2
Efeitos da mefedrona nas reduções do DAT estriatal induzidas pela metanfetamina. Os ratos foram tratados com as doses indicadas de mefedrona (MEPH) 30 min antes de cada injeção de 2.5 (●) ou 5.0 mg / kg (■) de metanfetamina (METH) e sacrificados 2d mais tarde para determinação dos níveis estriatais de DAT por immunoblotting (a). Os blots foram quantificados usando ImageJ e os dados são médios ± SEM para 10-12 ratos por grupo (b). *p <0.01 ou ***p <0.0001 vs controlo (C) e #p <0.01 ou ##p <0.001 vs a respectiva dose de metanfetamina (teste de comparação múltipla de Tukey).
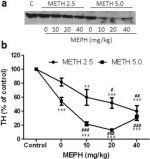
A Fig. 3a mostra que a mefedrona aumentou significativamente as reduções induzidas pela metanfetamina nos níveis de TH, conforme determinado por immunoblotting. Os immunoblots foram quantificados e, de acordo com os resultados acima para DA e DAT, os principais efeitos da dose de metanfetamina (F1,81 = 47,89, p <0,0001) e da dose de mefedrona (F4,81 = 63,57, p <0,0001) foram altamente significativos por ANOVA de duas vias (Fig. 3b). O efeito principal da mefedrona administrada em combinação com 2,5 mg/kg (F4,34 = 12,98, p < 0,0001) ou 5,0 mg/kg de metanfetamina (F4,49 = 99,16, p < 0,0001) também foi altamente significativo pela ANOVA de uma via. Todos os tratamentos com qualquer uma das doses de metanfetamina ± mefedrona causaram reduções significativamente maiores de TH em comparação com o respetivo controlo (p < 0,001 para 2,5 mg/kg de metanfetamina + 10 mg/kg de mefedrona; p < 0,0001 para todas as outras combinações), com exceção de 2,5 mg/kg de metanfetamina isolada, que não alterou significativamente os níveis de TH (ou seja, sem toxicidade). A figura 3b mostra igualmente que as doses de mefedrona de 20 mg/kg (p < 0,01) e 40 mg/kg (p < 0,001) aumentaram significativamente as reduções de TH causadas por 2,5 mg/kg de metanfetamina e que as três doses de mefedrona aumentaram significativamente (p < 0,0001) os efeitos de 5,0 mg/kg de metanfetamina nas reduções de TH.
Fig. 3
Efeitos da mefedrona nas reduções da TH estriatal induzidas pela metanfetamina. Os ratos foram tratados com as doses indicadas de mefedrona (MEPH) 30 min antes de cada injeção de 2.5 (●) ou 5.0 mg / kg (■) de metanfetamina (METH) e sacrificados 2d mais tarde para determinação dos níveis estriatais de TH por immunoblotting (a). Os blots foram quantificados usando ImageJ e os dados são médios ± SEM para 10-12 ratos por grupo (b). Algumas barras de erro eram demasiado pequenas para exceder o tamanho dos símbolos e não aparecem visíveis. **p < 0.001 ou ***p < 0.0001 vs controlo (C) e #p < 0.01, ##p < 0.001 ou ###p < 0.0001) vs a respectiva dose de metanfetamina (teste de comparação múltipla de Tukey).
Efeitos da mefedrona na hipertermia induzida pela metanfetamina
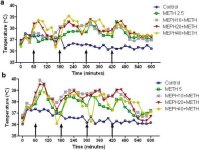
A mefedrona, tal como a metanfetamina, provoca uma hipertermia significativa (Hadlock et al. 2011, Baumann et al. 2012, Angoa-Perez et al. 2012). Quando a mefedrona foi administrada 30 minutos antes de cada injeção de metanfetamina, pode ser visto na Fig. 4 que os principais efeitos das doses de metanfetamina e mefedrona (F1,300 = 11.99, p <0.0001) ao longo do tempo (F4,300 = 51.73, p <0.0001) foram altamente significativos por ANOVA de duas vias. Os principais efeitos da mefedrona administrada em combinação com 2.5 mg / kg de metanfetamina (F4,120 = 41.44, p <0.0001, painel a) ao longo do tempo (F30,120 = 3.84, p <0.0001) ou 5.0 mg / kg de metanfetamina (F4,120 = 78.09, p <0.0001, painel b) ao longo do tempo (F30,120 = 9.98, p <0.0001) também foram altamente significativos pela ANOVA de duas vias. Todos os tratamentos com qualquer dose de metanfetamina ± mefedrona foram significativamente diferentes dos respectivos controlos (p <0,0001 para todos os tratamentos).
Fig. 4
Efeitos da mefedrona na hipertermia induzida pela metanfetamina. Os ratinhos foram tratados com as doses indicadas de mefedrona (MEPH) 30 minutos antes de cada injeção de 2,5 (a) ou 5,0 mg/kg (b) de metanfetamina (METH). As temperaturas centrais foram medidas a intervalos de 20 minutos por telemetria, com início 60 minutos antes da primeira injeção de metanfetamina. As 4 injecções de metanfetamina são indicadas pelas setas que repousam no eixo x. Os dados são expressos como temperatura corporal média de 6-8 ratos por grupo. Os SEMs foram sempre < 10% da média e são omitidos por uma questão de clareza.
Efeitos da mefedrona na neurotoxicidade induzida pela anfetamina e pelo MDMA
A fim de testar se os efeitos potenciadores da mefedrona na metanfetamina poderiam ser alargados a outras anfetaminas neurotóxicas, os ratinhos foram tratados com esta β-cetoanfetamina (20 mg/kg) mais anfetamina (4X 5 mg/kg) ou MDMA (4X 20 mg/kg) e os resultados são apresentados na Fig. 5. Recorde-se que a mefedrona em si não reduz a DA estriatal, a DAT ou a TH (Angoa-Perez et al. 2012). O principal efeito da droga (F5,27 = 27.18, p <0.0001) foi altamente significativo pela ANOVA unidirecional para reduções de DA (Fig. 5a). Também pode ser visto na Fig. 5a que todos os tratamentos com anfetamina (p <0.001) ou MDMA (p <0.001) sozinho ou em combinação com mefedrona (p <0.0001 para ambas as drogas) reduziram significativamente os níveis de DA do controle. A mefedrona aumentou significativamente as reduções de DA causadas por anfetamina (p <0.01) ou MDMA (p <0.01). Fig. 5b mostra efeitos semelhantes de tratamentos com drogas combinadas nos níveis de DAT no estriado. O efeito principal da droga (F4,49 = 42.63, p <0.0001) foi altamente significativo por ANOVA unidirecional para DAT. Também pode ser visto na Fig. 5b que todos os tratamentos com anfetamina ou MDMA foram significativamente (p <0.0001 para todos) mais baixos em comparação com o controle. A mefedrona também aumentou significativamente as reduções de DAT causadas por anfetamina ou MDMA (p <0.0001 em ambos os casos). Finalmente, a Fig. 5c mostra que o efeito principal da droga (F4,50 = 75.06, p <0.0001) foi altamente significativo por ANOVA unidirecional para reduções no TH. Também se pode ver na Fig. 5c que todos os tratamentos com anfetamina ou MDMA foram significativamente (p < 0.0001 para todos) mais baixos em comparação com o controlo. A mefedrona também aumentou significativamente as reduções de TH causadas pela anfetamina ou pelo MDMA (p < 0,0001 em ambos os casos)
Fig. 5
Efeitos da mefedrona na neurotoxicidade da terminação nervosa DA induzida por anfetamina ou MDMA. Os ratinhos foram tratados com 20 mg/kg de mefedrona (MEPH) 30 minutos antes de cada injeção de 5,0 mg/kg de anfetamina (AMPH) ou 20 mg/kg de MDMA e sacrificados 2d após o tratamento para determinação dos níveis estriatais de (a) DA por HPLC. (b) DAT e (c) TH foram determinados por immunoblotting e os blots foram quantificados utilizando ImageJ. Os immunoblots representativos para DAT e TH são incluídos como inserções nos painéis (b) e (c), respetivamente, e os tratamentos para ambos os painéis são indicados por 1,5: controlo; 2,6: MEPH; 3: AMPH; 4: AMPH + MEPH; 7: MDMA; e 8: MDMA + MEPH. Os dados são a média ± SEM para 5-12 ratos em cada grupo. **p < 0,001 ou ***p < 0,0001 vs controlo e #p < 0,01 ou ###p < 0,0001 vs AMPH ou MDMA (teste de comparação múltipla de Tukey).
Efeitos da nomifensina na neurotoxicidade induzida pela metanfetamina
A nomifensina, um potente bloqueador do DAT sem abuso conhecido nem potencial neurotóxico, foi testada quanto à sua capacidade de proteção contra a neurotoxicidade induzida pela metanfetamina e quanto ao contraste com as acções da mefedrona na toxicidade para as terminações nervosas DA causada pela metanfetamina, anfetamina e MDMA. Os resultados da Fig. 6a mostram que o efeito principal do fármaco (F3,16 = 63,39, p < 0,0001) nos níveis de DA foi altamente significativo na ANOVA unidirecional. A nomifensina sozinha não alterou os níveis de DA, mas a redução causada pela metanfetamina (p <0.0001) foi ligeiramente, mas significativamente revertida pela nomifensina (p <0.01). O efeito principal da droga (F3,20 = 16.78, p <0.0001) nos níveis de DAT foi altamente significativo pela ANOVA unidirecional, conforme mostrado na Fig. 6b. A nomifensina não alterou os níveis de DAT, mas forneceu proteção significativa (p <0.001) contra a redução no DAT estriatal causada pela metanfetamina (p <0.0001) em comparação com o controle. Finalmente, a Fig. 6c mostra que o efeito principal da droga (F3,15 = 14.10, p <0.0001) nos níveis de TH foi altamente significativo pela ANOVA unidirecional. Tal como se verificou para a DA e o DAT, a redução do TH causada pela metanfetamina (p < 0,0001) foi ligeiramente mas significativamente evitada pela nomifensina (p < 0,01).
Fig. 6
Efeitos da nomifensina na neurotoxicidade da terminação nervosa DA induzida pela metanfetamina. Os ratinhos foram tratados com 5,0 mg/kg de nomifensina (NOM) 30 minutos antes de cada injeção de 5,0 mg/kg de metanfetamina (METH) e sacrificados 2 dias mais tarde para determinação dos níveis estriatais de (a) DA por HPLC. (b) DAT e (c) TH foram determinados por immunoblotting e os blots foram quantificados utilizando ImageJ. Os immunoblots representativos para DAT e TH estão incluídos como inserções nos painéis (b) e (c) respetivamente. Os dados são a média mais SEM para 5-7 ratos por grupo. ***p < 0.0001 vs controlo (C) e #p < 0.01 ou ##p < 0.001 vs metanfetamina isolada (teste de comparação múltipla de Tukey).
Discussão
O objetivo do presente estudo foi determinar se a mefedrona impediria a toxicidade da terminação nervosa DA causada pela metanfetamina. Com base na sua semelhança química com a metanfetamina e a MDMA, esperava-se inicialmente que a mefedrona exercesse efeitos nocivos no sistema neuronal DA. No entanto, vários estudos estabeleceram quase simultaneamente que a mefedrona não era tóxica para as terminações nervosas DA (Angoa-Perez et al. 2012, Baumann et al. 2012, Hadlock et al. 2011). A questão de saber se esta droga causa danos ao sistema neuronal 5-HT permanece em aberto. Um estudo relatou reduções persistentes na função de terminação nervosa 5-HT (Hadlock et al. 2011), enquanto outro descobriu que a mefedrona não causou danos (Baumann et al. 2012). A mefedrona interage com a terminação nervosa DA de uma forma que sugere que, de facto, estimula a libertação e bloqueia a recaptação de DA através das suas interacções com o DAT. Uma faceta fundamental do mecanismo de ação neurotóxico da metanfetamina é a sua capacidade de aceder às terminações nervosas DA através do DAT e perturbar a homeostase da DA (Sulzer 2011). Se este passo inicial na cascata neurotóxica da metanfetamina for impedido pela inibição do DAT, a toxicidade é evitada (Pu et al. 1994, Poth et al. 2012, Marek et al. 1990, Schmidt e Gibb 1985). Pensámos que a mefedrona poderia ter esta mesma propriedade protetora que outros inibidores da DAT, mas observámos, em vez disso, um aumento significativo da toxicidade. Esta interação foi observada utilizando duas doses diferentes de metanfetamina que causam danos moderados ou graves nas terminações nervosas DA (4X 2,5 ou 5,0 mg/kg, respetivamente). Este efeito potenciador da mefedrona não se limitou à metanfetamina e estendeu-se à anfetamina e à MDMA, duas drogas que são frequentemente consumidas em conjunto com a mefedrona e outras β-cetoanfetaminas (Feyissa e Kelly 2008, Schifano et al. 2011, Kelly 2011). Por conseguinte, apesar de a mefedrona não causar toxicidade pelo menos nas terminações nervosas DA do estriado, potencia os efeitos neurotóxicos de outras drogas de abuso. Esta nova descoberta deve tornar o abuso de mefedrona ainda mais evidente, uma vez que a sua falta de neurotoxicidade intrínseca pode fazer com que pareça inócua.
A hipertermia é um efeito adverso agudo comummente notificado da ingestão de metanfetamina (Greene et al. 2008) e de β-cetoanfetamina em seres humanos (Borek e Holstege 2012, Prosser e Nelson 2012). Tal como a metanfetamina, muitas das drogas β-cetoanfetaminas também causam elevações significativas da temperatura central em roedores (Angoa-Perez et al. 2012, Hadlock et al. 2011, Baumann et al. 2012, Rockhold et al. 1997). Embora a hipertermia causada pela metanfetamina possa contribuir para os seus efeitos morfológicos e neuronais prejudiciais, não é necessariamente o caso de a hipertermia ser a causa direta destes efeitos (Kiyatkin e Sharma 2009). Registámos as temperaturas corporais centrais em ratos tratados com mefedrona e metanfetamina e observámos que o tratamento combinado não aumentou as temperaturas para além dos aumentos máximos observados após a administração isolada de qualquer uma das drogas. A metanfetamina causou um aumento da temperatura corporal relacionado com a dose e esta hipertermia foi invariável ao longo de toda a gama de doses de mefedrona testadas. De facto, a queda da temperatura corporal pós-injeção observada após o tratamento com mefedrona (Angoa-Perez et al. 2012) manteve-se com doses mais elevadas de mefedrona mais metanfetamina. Embora a hipertermia induzida pela droga não tenha sido reforçada pelo tratamento combinado, os efeitos neurotóxicos foram aditivos. Por conseguinte, pelo menos no caso presente, parece que os efeitos neurotóxicos da metanfetamina podem ser reforçados pela mefedrona de uma forma que é independente da hipertermia.
A mefedrona inibe claramente a função DAT e bloqueia a recaptação de DA in vitro (Lopez-Arnau et al. 2012, Hadlock et al. 2011, Kehr et al. 2011, Martinez-Clemente et al. 2012, Cozzi et al. 1999). A mefedrona desloca o WIN-35,428 do seu sítio de ligação no DAT, sugerindo que é um inibidor competitivo da captação de DA (Martinez-Clemente et al. 2012, Lopez-Arnau et al. 2012). A potência da mefedrona a este respeito é muito semelhante à da metanfetamina (Cozzi et al. 1999) e da MDMA (Escubedo et al. 2011). Não se sabe se a mefedrona é transportada pelo DAT, mas a metcatinona sim (Cozzi e Foley 2003). A nomifensina e o ácido anfonélico, que se ligam ao DAT e inibem a captação de DA, proporcionam uma proteção substancial contra a neurotoxicidade induzida pela metanfetamina (Pu et al. 1994, Marek et al. 1990, Schmidt e Gibb 1985, Poth et al. 2012) e os ratinhos que não possuem o DAT são resistentes à toxicidade neuronal da metanfetamina (Fumagalli et al. 1998). O facto de a mefedrona não ser neurotóxica e ser um bloqueador do DAT leva a prever que deve prevenir a toxicidade. Testámos a nomifensina a este respeito como controlo positivo e confirmámos que protege contra a depleção de DA, DAT e TH induzida pela metanfetamina. A nomifensina também inibe o transportador de norepinefrina (Brogden et al. 1979), mas essa propriedade não pode explicar os presentes resultados porque a maioria das β-cetoanfetaminas, incluindo a mefedrona, inibe o transportador de norepinefrina e bloqueia a captação de norepinefrina (Kelly 2011, Rothman et al. 2003, Cozzi et al. 1999, Sogawa et al. 2011, Lopez-Arnau et al. 2012). Um papel para o sistema neuronal 5-HT em algumas das ações farmacológicas da mefedrona é possível à luz da capacidade desta droga, como MDMA (Yamamoto et al. 1995), para causar efluxo de DA estriatal através de suas interações com receptores 5-HT2A (Lopez-Arnau et al. 2012, Martinez-Clemente et al. 2012). A hiper-locomoção causada pela mefedrona é dependente da 5-HT endógena (Lopez-Arnau et al. 2012) e esta droga também estimula a libertação de 5-HT e inibe a sua captação in vitro (Sogawa et al. 2011, Cozzi et al. 1999, Nagai et al. 2007, Hadlock et al. 2011, Lopez-Arnau et al. 2012, Martinez-Clemente et al. 2012) e in vivo (Baumann et al. 2012, Kehr et al. 2011). No entanto, podemos excluir um papel para a 5-HT endógena na neurotoxicidade DA de, pelo menos, metanfetamina, mostrando que os ratos geneticamente depletados de 5-HT mantêm a sua sensibilidade à neurotoxicidade (Thomas et al. 2010).
A mefedrona poderia aumentar a neurotoxicidade da metanfetamina através de vários mecanismos possíveis. Em primeiro lugar, a mefedrona poderia interagir com o VMAT para provocar a fuga de DA para o citoplasma da terminação nervosa pré-sináptica. Os tratamentos que aumentam o pool citoplasmático (ou seja, libertável pelo fármaco) de DA aumentam a neurotoxicidade da metanfetamina (Thomas et al. 2008, Thomas et al. 2009, Schmidt et al. 1985). Este mecanismo não é provável porque a metcatinona interage apenas fracamente com o VMAT (Cozzi et al. 1999). Em segundo lugar, a combinação de mefedrona e metanfetamina poderia ter um efeito sinérgico na libertação não-vesicular de DA, mas esta possibilidade também parece improvável à luz dos resultados que mostram que o tratamento de células CHO que expressam DAT ou SERT com metilona e metanfetamina não tem um efeito aditivo na libertação de DA ou 5-HT (Sogawa et al. 2011). Em terceiro lugar, a mefedrona pode interagir com o DAT de uma forma inovadora que contribui para a toxicidade aditiva. Foi demonstrado que a metilona em combinação com a metanfetamina causa uma citotoxicidade sinérgica em células CHO que expressam o DAT ou o SERT, mas não em células CHO de tipo selvagem sem os transportadores (Sogawa et al. 2011). A citotoxicidade observada em células de cultura nestes estudos (i.e., libertação de LDH) é muito diferente da lesão das terminações nervosas DA causada pela metanfetamina, mas este mecanismo sugere um papel interessante mas indefinido para o DAT na citotoxicidade acrescida. Por último, a mefedrona pode alterar o metabolismo da metanfetamina. A mefedrona é principalmente metabolizada por N-desmetilação (Meyer e Maurer 2010), tal como a metanfetamina e a MDMA (Caldwell 1976). O apoio a este mecanismo vem da demonstração de que a metanfetamina e o MDMA inibem mutuamente a produção dos seus respetivos metabolitos primários e elevam os níveis plasmáticos da droga acima dos observados após a administração de qualquer uma das drogas isoladamente (Kuwayama et al. 2012). As doses de mefedrona utilizadas atualmente e no nosso estudo anterior (Angoa-Perez et al. 2012), embora elevadas, não são neurotóxicas e situam-se no intervalo de consumo dos seres humanos (McErath e O'Neill 2011). Por conseguinte, a mefedrona pode estar a agir como a MDMA para aumentar os níveis plasmáticos de metanfetamina, inibindo o seu metabolismo. Será necessária uma análise farmacocinética aprofundada para confirmar esta última possibilidade.
O abuso das β-cetoanfetaminas está a aumentar a um ritmo alarmante e a mefedrona é atualmente uma das drogas mais consumidas, a seguir à canábis, à MDMA e à cocaína (Morris 2010, Winstock et al. 2011b). Além disso, a mefedrona induz sentimentos de desejo mais fortes nos seres humanos do que a MDMA (Brunt et al. 2011) e os utilizadores que snifam a mefedrona consideram-na mais viciante do que a cocaína (Winstock et al. 2011b). A mefedrona é consumida por seres humanos de forma compulsiva (ou seja, "stacking") e é frequentemente consumida com outras drogas, como a canábis e os psicoestimulantes anfetamínicos (Schifano et al. 2011, Fass et al. 2012, Winstock et al. 2011a, Kelly 2011, Torrance e Cooper 2010). A mefedrona é cada vez mais encontrada em comprimidos vendidos como MDMA (Brunt et al. 2011) e o seu consumo irá provavelmente ultrapassar o da MDMA, uma vez que a pureza desta última droga continua a diminuir (Brunt et al. 2011, Tanner-Smith 2006, Teng et al. 2006). Com base nos padrões comuns de abuso da mefedrona e de outros ingredientes dos "sais de banho", é importante considerar se existem riscos adicionais para a saúde dos seres humanos quando estas drogas são combinadas com as anfetaminas, propositada ou involuntariamente. Os nossos resultados, que mostram que pelo menos a mefedrona aumenta significativamente a neurotoxicidade das terminações nervosas DA do estriado causada pela metanfetamina, anfetamina e MDMA, revelam uma propriedade particularmente perigosa e inesperada desta β-cetoanfetamina.
Abreviaturas utilizadas
5-HT serotonina
DA dopamina
DAT transportador de DA
MDMA 3,4-metilenodioximetanfetamina
TH tirosina hidroxilase
VMAT transportador vesicular de monoamina
A mefedrona (4-metilmetcatinona) é um derivado da catinona e análogo estrutural da metanfetamina e da 3,4-metilenodioxi-metanfetamina (MDMA). A mefedrona é um dos ingredientes psicoactivos dos "sais de banho", juntamente com outros compostos, como a metilona, a butilona e a 3,4-metilenodioxipirovalerona (MDPV). As β-cetoanfetaminas estão a ser consumidas a taxas crescentes devido, em grande parte, à disponibilidade altamente restrita dos precursores necessários para a síntese de metanfetamina e MDMA em laboratórios clandestinos e à correspondente redução da sua pureza (Winstock et al. 2011b, Brunt et al. 2011). À medida que o abuso de β-cetoanfetaminas continua a aumentar, a lista dos seus efeitos adversos cresceu para incluir complicações cardiovasculares, agitação, insónia, psicose e depressão (Schifano et al. 2011, Prosser e Nelson 2012).
Como congéneres químicos da metanfetamina e da MDMA, não é surpreendente que as β-cetoanfetaminas tenham muitos dos mesmos efeitos que estas drogas anteriores no sistema nervoso central. Por exemplo, estas drogas bloqueiam os transportadores de dopamina (DA) e de serotonina (5-HT) (DAT e SERT, respetivamente) (Cozzi et al. 1999, Rothman et al. 2003, Fleckenstein et al. 2000, Lopez-Arnau et al. 2012) e estimulam a libertação de monoaminas in vitro (Kalix e Glennon 1986, Gygi et al. 1997, Rothman et al. 2003) e in vivo (Gygi et al. 1997, Kehr et al. 2011). A metcatinona provoca reduções persistentes da atividade da triptofano hidroxilase e da tirosina hidroxilase (TH) e depleção de DA e 5-HT (Gygi et al. 1997, Gygi et al. 1996, Sparago et al. 1996). Estudos de imagem PET em consumidores abstinentes de metcatinona revelaram uma redução da densidade do DAT no estriado, o que sugere uma perda de terminais DA (McCann et al. 1998). A estimulação simultânea da libertação de DA e a inibição da sua captação espelham os elementos críticos subjacentes à neurotoxicidade associada à metanfetamina (Kuhn et al. 2008, Yamamoto e Bankson 2005, Cadet et al. 2007, Fleckenstein et al. 2007).
Nós (Angoa-Perez et al. 2012) e outros (Baumann et al. 2012, Hadlock et al. 2011) investigámos recentemente a possibilidade de a mefedrona poder causar neurotoxicidade como a metanfetamina e a MDMA. Surpreendentemente, a mefedrona não era tóxica para as terminações nervosas DA do estriado (Hadlock et al. 2011, Baumann et al. 2012, Angoa-Perez et al. 2012). A questão de saber se a mefedrona danifica as terminações nervosas 5-HT continua por resolver, uma vez que um estudo documentou efeitos positivos (Hadlock et al. 2011), enquanto outro foi negativo (Baumann et al. 2012). À luz do efeito relativamente benigno da mefedrona nas terminações nervosas DA e considerando as suas propriedades como bloqueador DAT, colocámos a hipótese de que poderia realmente proteger o sistema neuronal DA dos efeitos neurotóxicos da metanfetamina, tal como se sabe que ocorre com outros bloqueadores DAT, como o ácido anfonélico (Pu et al. 1994, Schmidt e Gibb 1985, Marek et al. 1990) e nomifensina (Poth et al. 2012). Atualmente, verificamos que a mefedrona aumenta significativamente a neurotoxicidade da metanfetamina. Este efeito estende-se à anfetamina e à MDMA, drogas que são frequentemente consumidas em conjunto com a mefedrona (Feyissa e Kelly 2008, Schifano et al. 2011). Estes resultados surpreendentes lançam uma nova luz sobre o abuso da mefedrona e tornam urgente o reconhecimento desta propriedade subtil e perigosa desta β-cetoanfetamina.
Materiais e métodos
Drogas e reagentes
O cloridrato de mefedrona e a 3,4-metilenodioximetanfetamina (MDMA) foram obtidos no Programa de Fornecimento de Drogas do NIDA Research Resources. (+) O cloridrato de metanfetamina, o maleato de nomifensina, o sulfato de d-anfetamina, o pentobarbital, a DA e todos os tampões e reagentes de HPLC foram adquiridos à Sigma-Aldrich (St. Louis, MO, EUA). Os kits de ensaio de proteínas de ácido bicinconínico foram obtidos da Pierce (Rockford, IL, EUA). Os anticorpos policlonais contra a TH do rato foram produzidos conforme descrito anteriormente (Kuhn e Billingsley 1987). Os anticorpos monoclonais contra a DAT do rato foram generosamente fornecidos pela Dra. Roxanne A. Vaughan (Universidade do Dakota do Norte, Grand Forks, ND, EUA). Os anticorpos secundários anti-IgG conjugados com HRP foram fornecidos por Jackson ImmunoResearch Laboratories, Inc. (West Grove, PA, EUA).
Animais
As fêmeas de ratinhos C57BL/6 (Harlan, Indianapolis, IN, EUA), com um peso de 20-25 g na altura da experiência, foram alojadas 5 por gaiola em gaiolas grandes com caixa de sapatos, numa sala com luz (12 h de luz/obscuridade) e temperatura controlada. Foram utilizadas ratinhas fêmeas porque se sabe que são muito sensíveis aos danos neuronais provocados pelas anfetaminas neurotóxicas e para manter a coerência com os nossos estudos anteriores sobre a neurotoxicidade da metanfetamina (Thomas et al. 2010, Thomas et al. 2008, Thomas et al. 2009). Os ratinhos tinham livre acesso a alimentos e água. O Comité Institucional de Cuidados e Utilização da Wayne State University aprovou os cuidados com os animais e os procedimentos experimentais. Todos os procedimentos também estavam em conformidade com o Guia do NIH para o Cuidado e Uso de Animais de Laboratório.
Procedimentos farmacológicos, fisiológicos e comportamentais
Os ratinhos foram tratados com mefedrona utilizando um regime de tratamento em "binge" composto por 4 injecções de 10, 20 ou 40 mg/kg com um intervalo de 2 horas entre cada injeção. Este regime de tratamento em "binge", quando utilizado para injetar anfetaminas substituídas e derivados da catinona, provoca lesões extensas nas terminações nervosas DA. As doses de mefedrona atualmente utilizadas demonstraram anteriormente não serem tóxicas para as terminações nervosas DA (Angoa-Perez et al. 2012). Os ratinhos foram tratados com metanfetamina (4X 2,5 ou 5 mg/kg), anfetamina (4X 5 mg/kg) ou MDMA (4X 20 mg/kg) isoladamente ou em combinação com mefedrona. Quando tratados com duas drogas, os ratos receberam uma injeção de mefedrona 30 minutos antes de cada uma das 4 injecções de metanfetamina, anfetamina ou MDMA. Os controlos receberam injecções de soro fisiológico no mesmo horário utilizado para a mefedrona isolada ou em combinação com outras anfetaminas. Como controlo dos efeitos de um inibidor do DAT na toxicidade da metanfetamina, os ratinhos foram tratados com nomifensina (4X 5 mg/kg) 30 minutos antes de cada injeção de metanfetamina (4X 5 mg/kg). Todas as injecções foram administradas por via i.p.. Os ratinhos foram sacrificados 2 dias após o último tratamento medicamentoso, quando a neurotoxicidade associada à anfetamina atingiu o seu máximo. A temperatura corporal foi monitorizada por telemetria utilizando transponders de temperatura implantáveis IPTT-300 da Bio Medic Data Systems, Inc. (Seaford, DE, EUA). As temperaturas foram registadas de forma não invasiva a cada 20 minutos, começando 60 minutos antes da primeira injeção de METH e continuando durante as 9 horas seguintes, utilizando o sistema de consola DAS-5001 da Bio Medic.
Determinação do teor de DA no estriado
O tecido estriado foi dissecado bilateralmente do cérebro após o tratamento e armazenado a -80°C. Os tecidos congelados foram pesados e submetidos a sonicação em 10 volumes de ácido perclórico 0,16 N a 4°C. A proteína insolúvel foi removida por centrifugação e a DA foi determinada por HPLC com deteção eletroquímica, tal como descrito anteriormente para a metanfetamina (Thomas et al. 2010, Thomas et al, 2009).
Determinação dos níveis de proteínas TH e DAT por immunoblotting
Os efeitos dos tratamentos medicamentosos nos níveis estriatais de TH e DAT foram determinados por immunoblotting como um índice de toxicidade para as terminações nervosas DA estriatais. Os ratos foram sacrificados por decapitação após o tratamento e o estriado foi dissecado bilateralmente. O tecido foi armazenado a -80°C. O tecido congelado foi desfeito por sonicação em SDS a 1% a 95°C e o material insolúvel foi sedimentado por centrifugação. A proteína foi determinada pelo método do ácido bicinchonínico e quantidades iguais de proteína (70 μg/lano) foram resolvidas por eletroforese em gel de SDS-poliacrilamida e depois electroblotadas para nitrocelulose. Os blots foram bloqueados em solução salina tamponada com Tris contendo Tween 20 (0,1% v/v) e 5% de leite seco magro durante 1 h à temperatura ambiente. Os anticorpos primários contra TH (1:1000) ou DAT (1:1000) foram adicionados aos blots e deixados a incubar durante 16 horas a 4°C. Os blots foram lavados 3 vezes em solução salina tamponada com Tris para remover os anticorpos que não reagiram e depois incubados com anticorpo secundário anti-IgG conjugado com HRP (1:4000) durante 1 h à temperatura ambiente. As bandas imunorreactivas foram visualizadas por quimioluminescência reforçada e as densidades relativas das bandas reactivas a TH e DAT foram determinadas por imagem com uma Kodak Image Station (Carestream Molecular Systems, Rochester, NY, EUA) e quantificadas utilizando o software ImageJ (NIH).
Análise dos dados
Foram efectuadas ANOVAs de duas vias para analisar os efeitos da dose de metanfetamina versus mefedrona na DA, DAT e TH. Os efeitos dos tratamentos com drogas no conteúdo estriatal de DA, TH e DAT foram testados quanto à significância por ANOVA de uma via seguida do teste de comparação múltipla de Tukey. Os resultados dos tratamentos medicamentosos na temperatura corporal central ao longo do tempo foram analisados usando uma ANOVA de duas vias seguida pelo teste de Bonferroni para determinar a significância das diferenças de temperatura em momentos individuais após o tratamento. As diferenças foram consideradas significativas se p < 0,05. Todas as análises estatísticas foram realizadas usando o GraphPad Prism versão 5.02 para Windows (GraphPad Software, San Diego, CA, EUA, www.graphpad.com).
Ir para:
Resultados
Efeitos da mefedrona na neurotoxicidade induzida pela metanfetamina
A mefedrona, em doses (10, 20 ou 40 mg/kg) que se sabe não causarem toxicidade na terminação nervosa DA (Angoa-Perez et al. 2012) foi administrada 30 minutos antes de cada injeção de metanfetamina. A metanfetamina foi administrada em doses que causam danos moderados (4X 2,5 mg/kg) ou graves (4X 5 mg/kg) nas terminações nervosas DA do estriado (Thomas et al. 2004, Thomas et al. 2010). Os resultados apresentados na Fig. 1 mostram que os principais efeitos da dose de metanfetamina (F1,40 = 66.60, p <0.0001) e da dose de mefedrona (F4,40 = 131.3, p <0.0001) nos níveis de DA no estriado foram altamente significativos pela ANOVA de duas vias. O efeito principal da mefedrona administrada em combinação com 2.5 mg / kg (F4,22 = 35.96, p <0.001) ou 5.0 mg / kg de metanfetamina (F4,17 = 953.9, p <0.0001) também foi altamente significativo por ANOVA unidirecional. Todos os tratamentos com qualquer dose de metanfetamina ± mefedrona causaram reduções significativamente maiores em DA em comparação com o respetivo controle (p <0.0001 para todos). A Fig. 1 mostra também que as doses de mefedrona de 20 (p < 0,01) e 40 mg/kg (p < 0,001) aumentaram significativamente os efeitos depletores de 2,5 mg/kg de metanfetamina na AD, enquanto todas as doses de mefedrona aumentaram significativamente os efeitos de 5,0 mg/kg de metanfetamina nos níveis de AD (p < 0,0001 para todos).
http://bbzzzsvqcrqtki6umym6itiixfhni37ybtt7mkbjyxn2pgllzxf2qgyd.onion/index.php?attachments/sywobkqrnd-jpg.47/&hash=f31155ada4e59f22a35f9110aaa6a7e5
Fig.1
Efeitos da mefedrona nas reduções da DA estriatal induzidas pela metanfetamina. Os ratos foram tratados com as doses indicadas de mefedrona (MEPH) 30 min antes de cada injeção de 2.5 (-) ou 5.0 mg / kg (■) de metanfetamina (METH) e sacrificados 2d mais tarde para determinação dos níveis estriatais de DA por HPLC. Os dados são médios ± SEM para 5-7 ratos por grupo. Algumas barras de erro eram demasiado pequenas para exceder o tamanho dos símbolos e não aparecem visíveis. ***p < 0,001 vs controlos e #p < 0,01, ##p < 0,001 ou ###p < 0,0001 vs a respectiva dose de metanfetamina (teste de comparação múltipla de Tukey).
Fig. 2a mostra que a mefedrona aumentou significativamente as reduções induzidas pela metanfetamina nos níveis de DAT, conforme determinado por immunoblotting. Os immunoblots foram quantificados e, de acordo com os resultados para DA, os principais efeitos da dose de metanfetamina (F1,92 = 9.48, p <0.001) e da dose de mefedrona (F4,92 = 37.56, p <0.0001) nos níveis de DAT no estriado foram altamente significativos por ANOVA de duas vias (Fig. 2b). O efeito principal da mefedrona administrada em combinação com 2.5 mg / kg (F4,56 = 15.55, p <0.0001) ou 5.0 mg / kg de metanfetamina (F4,39 = 24.84, p <0.0001) também foi altamente significativo por ANOVA unidirecional. Todos os tratamentos com qualquer dose de metanfetamina ± mefedrona causaram reduções significativamente maiores no DAT em comparação com o respetivo controle (p <0.01 para 2.5 mg / kg de metanfetamina sozinho; p <0.0001 para todos os outros tratamentos). A Fig. 2b mostra também que as doses de mefedrona de 20 mg/kg (p < 0,01) e 40 mg/kg (p < 0,001) aumentaram significativamente as reduções do DAT causadas por 2,5 mg/kg de metanfetamina, ao passo que apenas a dose de 40 mg/kg de mefedrona aumentou significativamente (p < 0,01) os efeitos de 5,0 mg/kg de metanfetamina nas reduções do DAT.
http://bbzzzsvqcrqtki6umym6itiixfhni37ybtt7mkbjyxn2pgllzxf2qgyd.onion/index.php?attachments/bfu3sx5dnr-jpg.4797/&hash=f31155ada4e59f22a35f9110aaa6a7e5
Fig.2
Efeitos da mefedrona nas reduções do DAT estriatal induzidas pela metanfetamina. Os ratos foram tratados com as doses indicadas de mefedrona (MEPH) 30 min antes de cada injeção de 2.5 (●) ou 5.0 mg / kg (■) de metanfetamina (METH) e sacrificados 2d mais tarde para determinação dos níveis estriatais de DAT por immunoblotting (a). Os blots foram quantificados usando ImageJ e os dados são médios ± SEM para 10-12 ratos por grupo (b). *p <0.01 ou ***p <0.0001 vs controlo (C) e #p <0.01 ou ##p <0.001 vs a respectiva dose de metanfetamina (teste de comparação múltipla de Tukey).
A Fig. 3a mostra que a mefedrona aumentou significativamente as reduções induzidas pela metanfetamina nos níveis de TH, conforme determinado por immunoblotting. Os immunoblots foram quantificados e, de acordo com os resultados acima para DA e DAT, os principais efeitos da dose de metanfetamina (F1,81 = 47,89, p <0,0001) e da dose de mefedrona (F4,81 = 63,57, p <0,0001) foram altamente significativos por ANOVA de duas vias (Fig. 3b). O efeito principal da mefedrona administrada em combinação com 2,5 mg/kg (F4,34 = 12,98, p < 0,0001) ou 5,0 mg/kg de metanfetamina (F4,49 = 99,16, p < 0,0001) também foi altamente significativo pela ANOVA de uma via. Todos os tratamentos com qualquer uma das doses de metanfetamina ± mefedrona causaram reduções significativamente maiores de TH em comparação com o respetivo controlo (p < 0,001 para 2,5 mg/kg de metanfetamina + 10 mg/kg de mefedrona; p < 0,0001 para todas as outras combinações), com exceção de 2,5 mg/kg de metanfetamina isolada, que não alterou significativamente os níveis de TH (ou seja, sem toxicidade). A figura 3b mostra igualmente que as doses de mefedrona de 20 mg/kg (p < 0,01) e 40 mg/kg (p < 0,001) aumentaram significativamente as reduções de TH causadas por 2,5 mg/kg de metanfetamina e que as três doses de mefedrona aumentaram significativamente (p < 0,0001) os efeitos de 5,0 mg/kg de metanfetamina nas reduções de TH.
http://bbzzzsvqcrqtki6umym6itiixfhni37ybtt7mkbjyxn2pgllzxf2qgyd.onion/index.php?attachments/tggfrjpqru-jpg.4798/&hash=f31155ada4e59f22a35f9110aaa6a7e5
Fig. 3
Efeitos da mefedrona nas reduções da TH estriatal induzidas pela metanfetamina. Os ratos foram tratados com as doses indicadas de mefedrona (MEPH) 30 min antes de cada injeção de 2.5 (●) ou 5.0 mg / kg (■) de metanfetamina (METH) e sacrificados 2d mais tarde para determinação dos níveis estriatais de TH por immunoblotting (a). Os blots foram quantificados usando ImageJ e os dados são médios ± SEM para 10-12 ratos por grupo (b). Algumas barras de erro eram demasiado pequenas para exceder o tamanho dos símbolos e não aparecem visíveis. **p < 0.001 ou ***p < 0.0001 vs controlo (C) e #p < 0.01, ##p < 0.001 ou ###p < 0.0001) vs a respectiva dose de metanfetamina (teste de comparação múltipla de Tukey).
Efeitos da mefedrona na hipertermia induzida pela metanfetamina
A mefedrona, tal como a metanfetamina, provoca uma hipertermia significativa (Hadlock et al. 2011, Baumann et al. 2012, Angoa-Perez et al. 2012). Quando a mefedrona foi administrada 30 minutos antes de cada injeção de metanfetamina, pode ser visto na Fig. 4 que os principais efeitos das doses de metanfetamina e mefedrona (F1,300 = 11.99, p <0.0001) ao longo do tempo (F4,300 = 51.73, p <0.0001) foram altamente significativos por ANOVA de duas vias. Os principais efeitos da mefedrona administrada em combinação com 2.5 mg / kg de metanfetamina (F4,120 = 41.44, p <0.0001, painel a) ao longo do tempo (F30,120 = 3.84, p <0.0001) ou 5.0 mg / kg de metanfetamina (F4,120 = 78.09, p <0.0001, painel b) ao longo do tempo (F30,120 = 9.98, p <0.0001) também foram altamente significativos pela ANOVA de duas vias. Todos os tratamentos com qualquer dose de metanfetamina ± mefedrona foram significativamente diferentes dos respectivos controlos (p <0,0001 para todos os tratamentos).
http://bbzzzsvqcrqtki6umym6itiixfhni37ybtt7mkbjyxn2pgllzxf2qgyd.onion/index.php?attachments/x0ik9hdwsw-jpg.4799/&hash=f31155ada4e59f22a35f9110aaa6a7e5
Fig. 4
Efeitos da mefedrona na hipertermia induzida pela metanfetamina. Os ratinhos foram tratados com as doses indicadas de mefedrona (MEPH) 30 minutos antes de cada injeção de 2,5 (a) ou 5,0 mg/kg (b) de metanfetamina (METH). As temperaturas centrais foram medidas a intervalos de 20 minutos por telemetria, com início 60 minutos antes da primeira injeção de metanfetamina. As 4 injecções de metanfetamina são indicadas pelas setas que repousam no eixo x. Os dados são expressos como temperatura corporal média de 6-8 ratos por grupo. Os SEMs foram sempre < 10% da média e são omitidos por uma questão de clareza.
Efeitos da mefedrona na neurotoxicidade induzida pela anfetamina e pelo MDMA
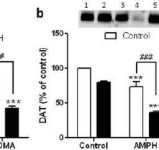
A fim de testar se os efeitos potenciadores da mefedrona na metanfetamina poderiam ser alargados a outras anfetaminas neurotóxicas, os ratinhos foram tratados com esta β-cetoanfetamina (20 mg/kg) mais anfetamina (4X 5 mg/kg) ou MDMA (4X 20 mg/kg) e os resultados são apresentados na Fig. 5. Recorde-se que a mefedrona em si não reduz a DA estriatal, a DAT ou a TH (Angoa-Perez et al. 2012). O principal efeito da droga (F5,27 = 27.18, p <0.0001) foi altamente significativo pela ANOVA unidirecional para reduções de DA (Fig. 5a). Também pode ser visto na Fig. 5a que todos os tratamentos com anfetamina (p <0.001) ou MDMA (p <0.001) sozinho ou em combinação com mefedrona (p <0.0001 para ambas as drogas) reduziram significativamente os níveis de DA do controle. A mefedrona aumentou significativamente as reduções de DA causadas por anfetamina (p <0.01) ou MDMA (p <0.01). Fig. 5b mostra efeitos semelhantes de tratamentos com drogas combinadas nos níveis de DAT no estriado. O efeito principal da droga (F4,49 = 42.63, p <0.0001) foi altamente significativo por ANOVA unidirecional para DAT. Também pode ser visto na Fig. 5b que todos os tratamentos com anfetamina ou MDMA foram significativamente (p <0.0001 para todos) mais baixos em comparação com o controle. A mefedrona também aumentou significativamente as reduções de DAT causadas por anfetamina ou MDMA (p <0.0001 em ambos os casos). Finalmente, a Fig. 5c mostra que o efeito principal da droga (F4,50 = 75.06, p <0.0001) foi altamente significativo por ANOVA unidirecional para reduções no TH. Também se pode ver na Fig. 5c que todos os tratamentos com anfetamina ou MDMA foram significativamente (p < 0.0001 para todos) mais baixos em comparação com o controlo. A mefedrona também aumentou significativamente as reduções de TH causadas pela anfetamina ou pelo MDMA (p < 0,0001 em ambos os casos)
http://bbzzzsvqcrqtki6umym6itiixfhni37ybtt7mkbjyxn2pgllzxf2qgyd.onion/index.php?attachments/onvypkgxvn-jpg.4800/&hash=f31155ada4e59f22a35f9110aaa6a7e5
Fig. 5
Efeitos da mefedrona na neurotoxicidade da terminação nervosa DA induzida por anfetamina ou MDMA. Os ratinhos foram tratados com 20 mg/kg de mefedrona (MEPH) 30 minutos antes de cada injeção de 5,0 mg/kg de anfetamina (AMPH) ou 20 mg/kg de MDMA e sacrificados 2d após o tratamento para determinação dos níveis estriatais de (a) DA por HPLC. (b) DAT e (c) TH foram determinados por immunoblotting e os blots foram quantificados utilizando ImageJ. Os immunoblots representativos para DAT e TH são incluídos como inserções nos painéis (b) e (c), respetivamente, e os tratamentos para ambos os painéis são indicados por 1,5: controlo; 2,6: MEPH; 3: AMPH; 4: AMPH + MEPH; 7: MDMA; e 8: MDMA + MEPH. Os dados são a média ± SEM para 5-12 ratos em cada grupo. **p < 0,001 ou ***p < 0,0001 vs controlo e #p < 0,01 ou ###p < 0,0001 vs AMPH ou MDMA (teste de comparação múltipla de Tukey).
Efeitos da nomifensina na neurotoxicidade induzida pela metanfetamina
A nomifensina, um potente bloqueador do DAT sem abuso conhecido nem potencial neurotóxico, foi testada quanto à sua capacidade de proteção contra a neurotoxicidade induzida pela metanfetamina e quanto ao contraste com as acções da mefedrona na toxicidade para as terminações nervosas DA causada pela metanfetamina, anfetamina e MDMA. Os resultados da Fig. 6a mostram que o efeito principal do fármaco (F3,16 = 63,39, p < 0,0001) nos níveis de DA foi altamente significativo na ANOVA unidirecional. A nomifensina sozinha não alterou os níveis de DA, mas a redução causada pela metanfetamina (p <0.0001) foi ligeiramente, mas significativamente revertida pela nomifensina (p <0.01). O efeito principal da droga (F3,20 = 16.78, p <0.0001) nos níveis de DAT foi altamente significativo pela ANOVA unidirecional, conforme mostrado na Fig. 6b. A nomifensina não alterou os níveis de DAT, mas forneceu proteção significativa (p <0.001) contra a redução no DAT estriatal causada pela metanfetamina (p <0.0001) em comparação com o controle. Finalmente, a Fig. 6c mostra que o efeito principal da droga (F3,15 = 14.10, p <0.0001) nos níveis de TH foi altamente significativo pela ANOVA unidirecional. Tal como se verificou para a DA e o DAT, a redução do TH causada pela metanfetamina (p < 0,0001) foi ligeiramente mas significativamente evitada pela nomifensina (p < 0,01).
http://bbzzzsvqcrqtki6umym6itiixfhni37ybtt7mkbjyxn2pgllzxf2qgyd.onion/index.php?attachments/daczg9cwfv-jpg.4801/&hash=f31155ada4e59f22a35f9110aaa6a7e5
Fig. 6
Efeitos da nomifensina na neurotoxicidade da terminação nervosa DA induzida pela metanfetamina. Os ratinhos foram tratados com 5,0 mg/kg de nomifensina (NOM) 30 minutos antes de cada injeção de 5,0 mg/kg de metanfetamina (METH) e sacrificados 2 dias mais tarde para determinação dos níveis estriatais de (a) DA por HPLC. (b) DAT e (c) TH foram determinados por immunoblotting e os blots foram quantificados utilizando ImageJ. Os immunoblots representativos para DAT e TH estão incluídos como inserções nos painéis (b) e (c) respetivamente. Os dados são a média mais SEM para 5-7 ratos por grupo. ***p < 0.0001 vs controlo (C) e #p < 0.01 ou ##p < 0.001 vs metanfetamina isolada (teste de comparação múltipla de Tukey).
Discussão
O objetivo do presente estudo foi determinar se a mefedrona impediria a toxicidade da terminação nervosa DA causada pela metanfetamina. Com base na sua semelhança química com a metanfetamina e a MDMA, esperava-se inicialmente que a mefedrona exercesse efeitos nocivos no sistema neuronal DA. No entanto, vários estudos estabeleceram quase simultaneamente que a mefedrona não era tóxica para as terminações nervosas DA (Angoa-Perez et al. 2012, Baumann et al. 2012, Hadlock et al. 2011). A questão de saber se esta droga causa danos ao sistema neuronal 5-HT permanece em aberto. Um estudo relatou reduções persistentes na função de terminação nervosa 5-HT (Hadlock et al. 2011), enquanto outro descobriu que a mefedrona não causou danos (Baumann et al. 2012). A mefedrona interage com a terminação nervosa DA de uma forma que sugere que, de facto, estimula a libertação e bloqueia a recaptação de DA através das suas interacções com o DAT. Uma faceta fundamental do mecanismo de ação neurotóxico da metanfetamina é a sua capacidade de aceder às terminações nervosas DA através do DAT e perturbar a homeostase da DA (Sulzer 2011). Se este passo inicial na cascata neurotóxica da metanfetamina for impedido pela inibição do DAT, a toxicidade é evitada (Pu et al. 1994, Poth et al. 2012, Marek et al. 1990, Schmidt e Gibb 1985). Pensámos que a mefedrona poderia ter esta mesma propriedade protetora que outros inibidores da DAT, mas observámos, em vez disso, um aumento significativo da toxicidade. Esta interação foi observada utilizando duas doses diferentes de metanfetamina que causam danos moderados ou graves nas terminações nervosas DA (4X 2,5 ou 5,0 mg/kg, respetivamente). Este efeito potenciador da mefedrona não se limitou à metanfetamina e estendeu-se à anfetamina e à MDMA, duas drogas que são frequentemente consumidas em conjunto com a mefedrona e outras β-cetoanfetaminas (Feyissa e Kelly 2008, Schifano et al. 2011, Kelly 2011). Por conseguinte, apesar de a mefedrona não causar toxicidade pelo menos nas terminações nervosas DA do estriado, potencia os efeitos neurotóxicos de outras drogas de abuso. Esta nova descoberta deve tornar o abuso de mefedrona ainda mais evidente, uma vez que a sua falta de neurotoxicidade intrínseca pode fazer com que pareça inócua.
A hipertermia é um efeito adverso agudo comummente notificado da ingestão de metanfetamina (Greene et al. 2008) e de β-cetoanfetamina em seres humanos (Borek e Holstege 2012, Prosser e Nelson 2012). Tal como a metanfetamina, muitas das drogas β-cetoanfetaminas também causam elevações significativas da temperatura central em roedores (Angoa-Perez et al. 2012, Hadlock et al. 2011, Baumann et al. 2012, Rockhold et al. 1997). Embora a hipertermia causada pela metanfetamina possa contribuir para os seus efeitos morfológicos e neuronais prejudiciais, não é necessariamente o caso de a hipertermia ser a causa direta destes efeitos (Kiyatkin e Sharma 2009). Registámos as temperaturas corporais centrais em ratos tratados com mefedrona e metanfetamina e observámos que o tratamento combinado não aumentou as temperaturas para além dos aumentos máximos observados após a administração isolada de qualquer uma das drogas. A metanfetamina causou um aumento da temperatura corporal relacionado com a dose e esta hipertermia foi invariável ao longo de toda a gama de doses de mefedrona testadas. De facto, a queda da temperatura corporal pós-injeção observada após o tratamento com mefedrona (Angoa-Perez et al. 2012) manteve-se com doses mais elevadas de mefedrona mais metanfetamina. Embora a hipertermia induzida pela droga não tenha sido reforçada pelo tratamento combinado, os efeitos neurotóxicos foram aditivos. Por conseguinte, pelo menos no caso presente, parece que os efeitos neurotóxicos da metanfetamina podem ser reforçados pela mefedrona de uma forma que é independente da hipertermia.
A mefedrona inibe claramente a função DAT e bloqueia a recaptação de DA in vitro (Lopez-Arnau et al. 2012, Hadlock et al. 2011, Kehr et al. 2011, Martinez-Clemente et al. 2012, Cozzi et al. 1999). A mefedrona desloca o WIN-35,428 do seu sítio de ligação no DAT, sugerindo que é um inibidor competitivo da captação de DA (Martinez-Clemente et al. 2012, Lopez-Arnau et al. 2012). A potência da mefedrona a este respeito é muito semelhante à da metanfetamina (Cozzi et al. 1999) e da MDMA (Escubedo et al. 2011). Não se sabe se a mefedrona é transportada pelo DAT, mas a metcatinona sim (Cozzi e Foley 2003). A nomifensina e o ácido anfonélico, que se ligam ao DAT e inibem a captação de DA, proporcionam uma proteção substancial contra a neurotoxicidade induzida pela metanfetamina (Pu et al. 1994, Marek et al. 1990, Schmidt e Gibb 1985, Poth et al. 2012) e os ratinhos que não possuem o DAT são resistentes à toxicidade neuronal da metanfetamina (Fumagalli et al. 1998). O facto de a mefedrona não ser neurotóxica e ser um bloqueador do DAT leva a prever que deve prevenir a toxicidade. Testámos a nomifensina a este respeito como controlo positivo e confirmámos que protege contra a depleção de DA, DAT e TH induzida pela metanfetamina. A nomifensina também inibe o transportador de norepinefrina (Brogden et al. 1979), mas essa propriedade não pode explicar os presentes resultados porque a maioria das β-cetoanfetaminas, incluindo a mefedrona, inibe o transportador de norepinefrina e bloqueia a captação de norepinefrina (Kelly 2011, Rothman et al. 2003, Cozzi et al. 1999, Sogawa et al. 2011, Lopez-Arnau et al. 2012). Um papel para o sistema neuronal 5-HT em algumas das ações farmacológicas da mefedrona é possível à luz da capacidade desta droga, como MDMA (Yamamoto et al. 1995), para causar efluxo de DA estriatal através de suas interações com receptores 5-HT2A (Lopez-Arnau et al. 2012, Martinez-Clemente et al. 2012). A hiper-locomoção causada pela mefedrona é dependente da 5-HT endógena (Lopez-Arnau et al. 2012) e esta droga também estimula a libertação de 5-HT e inibe a sua captação in vitro (Sogawa et al. 2011, Cozzi et al. 1999, Nagai et al. 2007, Hadlock et al. 2011, Lopez-Arnau et al. 2012, Martinez-Clemente et al. 2012) e in vivo (Baumann et al. 2012, Kehr et al. 2011). No entanto, podemos excluir um papel para a 5-HT endógena na neurotoxicidade DA de, pelo menos, metanfetamina, mostrando que os ratos geneticamente depletados de 5-HT mantêm a sua sensibilidade à neurotoxicidade (Thomas et al. 2010).
A mefedrona poderia aumentar a neurotoxicidade da metanfetamina através de vários mecanismos possíveis. Em primeiro lugar, a mefedrona poderia interagir com o VMAT para provocar a fuga de DA para o citoplasma da terminação nervosa pré-sináptica. Os tratamentos que aumentam o pool citoplasmático (ou seja, libertável pelo fármaco) de DA aumentam a neurotoxicidade da metanfetamina (Thomas et al. 2008, Thomas et al. 2009, Schmidt et al. 1985). Este mecanismo não é provável porque a metcatinona interage apenas fracamente com o VMAT (Cozzi et al. 1999). Em segundo lugar, a combinação de mefedrona e metanfetamina poderia ter um efeito sinérgico na libertação não-vesicular de DA, mas esta possibilidade também parece improvável à luz dos resultados que mostram que o tratamento de células CHO que expressam DAT ou SERT com metilona e metanfetamina não tem um efeito aditivo na libertação de DA ou 5-HT (Sogawa et al. 2011). Em terceiro lugar, a mefedrona pode interagir com o DAT de uma forma inovadora que contribui para a toxicidade aditiva. Foi demonstrado que a metilona em combinação com a metanfetamina causa uma citotoxicidade sinérgica em células CHO que expressam o DAT ou o SERT, mas não em células CHO de tipo selvagem sem os transportadores (Sogawa et al. 2011). A citotoxicidade observada em células de cultura nestes estudos (i.e., libertação de LDH) é muito diferente da lesão das terminações nervosas DA causada pela metanfetamina, mas este mecanismo sugere um papel interessante mas indefinido para o DAT na citotoxicidade acrescida. Por último, a mefedrona pode alterar o metabolismo da metanfetamina. A mefedrona é principalmente metabolizada por N-desmetilação (Meyer e Maurer 2010), tal como a metanfetamina e a MDMA (Caldwell 1976). O apoio a este mecanismo vem da demonstração de que a metanfetamina e o MDMA inibem mutuamente a produção dos seus respetivos metabolitos primários e elevam os níveis plasmáticos da droga acima dos observados após a administração de qualquer uma das drogas isoladamente (Kuwayama et al. 2012). As doses de mefedrona utilizadas atualmente e no nosso estudo anterior (Angoa-Perez et al. 2012), embora elevadas, não são neurotóxicas e situam-se no intervalo de consumo dos seres humanos (McErath e O'Neill 2011). Por conseguinte, a mefedrona pode estar a agir como a MDMA para aumentar os níveis plasmáticos de metanfetamina, inibindo o seu metabolismo. Será necessária uma análise farmacocinética aprofundada para confirmar esta última possibilidade.
O abuso das β-cetoanfetaminas está a aumentar a um ritmo alarmante e a mefedrona é atualmente uma das drogas mais consumidas, a seguir à canábis, à MDMA e à cocaína (Morris 2010, Winstock et al. 2011b). Além disso, a mefedrona induz sentimentos de desejo mais fortes nos seres humanos do que a MDMA (Brunt et al. 2011) e os utilizadores que snifam a mefedrona consideram-na mais viciante do que a cocaína (Winstock et al. 2011b). A mefedrona é consumida por seres humanos de forma compulsiva (ou seja, "stacking") e é frequentemente consumida com outras drogas, como a canábis e os psicoestimulantes anfetamínicos (Schifano et al. 2011, Fass et al. 2012, Winstock et al. 2011a, Kelly 2011, Torrance e Cooper 2010). A mefedrona é cada vez mais encontrada em comprimidos vendidos como MDMA (Brunt et al. 2011) e o seu consumo irá provavelmente ultrapassar o da MDMA, uma vez que a pureza desta última droga continua a diminuir (Brunt et al. 2011, Tanner-Smith 2006, Teng et al. 2006). Com base nos padrões comuns de abuso da mefedrona e de outros ingredientes dos "sais de banho", é importante considerar se existem riscos adicionais para a saúde dos seres humanos quando estas drogas são combinadas com as anfetaminas, propositada ou involuntariamente. Os nossos resultados, que mostram que pelo menos a mefedrona aumenta significativamente a neurotoxicidade das terminações nervosas DA do estriado causada pela metanfetamina, anfetamina e MDMA, revelam uma propriedade particularmente perigosa e inesperada desta β-cetoanfetamina.
Abreviaturas utilizadas
5-HT serotonina
DA dopamina
DAT transportador de DA
MDMA 3,4-metilenodioximetanfetamina
TH tirosina hidroxilase
VMAT transportador vesicular de monoamina