Mefedron (4-metylmetkatinon) är en β-ketoamfetaminstimulerande missbruksdrog med nära strukturella och mekanistiska likheter med metamfetamin. En av de mest kraftfulla effekterna som förknippas med mefedron är förmågan att stimulera frisättning av dopamin (DA) och blockera dess återupptag genom dess interaktion med dopamintransportören (DAT). Även om mefedron inte orsakar toxicitet för DA-nervändningar, kan dess förmåga att fungera som en DAT-blockerare ge skydd mot metamfetamininducerad neurotoxicitet som andra DAT-hämmare. För att testa denna möjlighet behandlades möss med mefedron (10, 20 eller 40 mg/kg) före varje injektion av en neurotoxisk regim av metamfetamin (4 injektioner av 2,5 eller 5,0 mg/kg med 2 timmars intervall). Integriteten hos DA-nervändarna i striatum bedömdes genom mätningar av DA-, DAT- och tyrosinhydroxylasnivåer. Den måttliga till allvarliga DA-toxicitet som förknippades med de olika doserna av metamfetamin förhindrades inte av någon dos av mefedron utan förstärktes i själva verket signifikant. Den hypertermi som orsakades av kombinerad behandling med mefedron och metamfetamin var densamma som efter behandling med något av läkemedlen ensamt. Mefedron förstärkte också de neurotoxiska effekterna av amfetamin och MDMA på DA-nervändarna. Däremot skyddade nomifensin mot metamfetamininducerad neurotoxicitet. Eftersom mefedron ökar metamfetamins neurotoxicitet, tyder de nuvarande resultaten på att det interagerar med DAT på ett sätt som inte liknar andra typiska DAT-hämmare. De relativt oskadliga effekterna av mefedron ensam på DA-nervändningar maskerar en potentiellt farlig interaktion med läkemedel som ofta missbrukas tillsammans med det, vilket leder till ökad neurotoxicitet.
Mefedron (4-metylmetkatinon) är ett katinonderivat och en strukturell analog till metamfetamin och 3,4-metylendioximetamfetamin (MDMA). Mefedron är en psykoaktiv ingrediens i "badsalter" tillsammans med andra föreningar som metylon, butylon och 3,4-metylendioxypyrovaleron (MDPV). β-ketoamfetaminer missbrukas i allt högre grad, vilket till stor del beror på den mycket begränsade tillgången på prekursorer som behövs för syntes av metamfetamin och MDMA i hemliga laboratorier och en motsvarande minskning av deras renhet (Winstock m.fl. 2011b, Brunt m.fl. 2011). I takt med att missbruket av β-ketoamfetaminer fortsätter att öka har listan över deras negativa effekter utökats till att omfatta kardiovaskulära komplikationer, agitation, sömnlöshet, psykoser och depression (Schifano m.fl. 2011, Prosser och Nelson 2012).
Som kemiska släktingar till metamfetamin och MDMA är det inte förvånande att β-ketoamfetaminerna har många av samma effekter på det centrala nervsystemet som de förstnämnda drogerna. Till exempel blockerar dessa läkemedel dopamin- (DA) och serotonin- (5-HT) transportörer (DAT respektive SERT) (Cozzi et al. 1999, Rothman et al. 2003, Fleckenstein et al. 2000, Lopez-Arnau et al. 2012) och de stimulerar monoaminfrisättning in vitro (Kalix och Glennon 1986, Gygi et al. 1997, Rothman et al. 2003) och in vivo (Gygi et al. 1997, Kehr et al. 2011). Metkatinon orsakar ihållande minskningar av tryptofanhydroxylas- och tyrosinhydroxylas (TH)-aktivitet och utarmning av DA och 5-HT (Gygi et al. 1997, Gygi et al. 1996, Sparago et al. 1996). PET-avbildningsstudier hos avhållsamma metkatinonanvändare avslöjade minskad DAT-densitet i striatal, vilket tyder på en förlust av DA-terminaler (McCann et al. 1998). Den samtidiga stimuleringen av DA-frisättning och hämningen av dess upptag speglar de kritiska element som ligger bakom neurotoxiciteten i samband med metamfetamin (Kuhn et al. 2008, Yamamoto och Bankson 2005, Cadet et al. 2007, Fleckenstein et al. 2007).
Vi (Angoa-Perez et al. 2012) och andra (Baumann et al. 2012, Hadlock et al. 2011) har nyligen undersökt möjligheten att mefedron skulle kunna orsaka neurotoxicitet på samma sätt som metamfetamin och MDMA. Överraskande nog var mefedron inte giftigt för DA-nervändarna i striatum (Hadlock et al. 2011, Baumann et al. 2012, Angoa-Perez et al. 2012). Frågan om huruvida mefedron skadar 5-HT-nervändarna är fortfarande oklar eftersom en studie dokumenterade positiva effekter (Hadlock et al. 2011) medan en annan var negativ (Baumann et al. 2012). Mot bakgrund av den relativt godartade effekten av mefedron på DA-nervändar och med tanke på dess egenskaper som DAT-blockerare, antog vi att det faktiskt skulle kunna skydda DA-neuronsystemet från de neurotoxiska effekterna av metamfetamin, precis som det är känt att det förekommer med andra DAT-blockerare såsom amfonsyra (Pu et al. 1994, Schmidt och Gibb 1985, Marek et al. 1990) och nomifensin (Poth et al. 2012). Vi rapporterar för närvarande att mefedron avsevärt förstärker neurotoxiciteten hos metamfetamin. Denna effekt gäller även amfetamin och MDMA, droger som ofta missbrukas tillsammans med mefedron (Feyissa och Kelly 2008, Schifano et al. 2011). Dessa överraskande resultat sätter missbruket av mefedron i ett nytt ljus och gör det angeläget att uppmärksamma denna subtila och farliga egenskap hos denna β-ketoamfetamin.
Material och metoder
Läkemedel och reagenser
Mefedronhydroklorid och 3,4-metylendioxymetamfetamin (MDMA) erhölls från NIDA Research Resources Drug Supply Program. (+) Metamfetaminhydroklorid, nomifensinmaleat, d-amfetaminsulfat, pentobarbital, DA och alla buffertar och HPLC-reagenser köptes från Sigma-Aldrich (St. Louis, MO, USA). Bicinchoninic acid protein assay kit erhölls från Pierce (Rockford, IL, USA). Polyklonala antikroppar mot TH från råttor framställdes enligt tidigare beskrivning (Kuhn och Billingsley 1987). Monoklonala antikroppar mot DAT från råtta tillhandahölls generöst av Dr. Roxanne A. Vaughan (University of North Dakota, Grand Forks, ND, USA). HRP-konjugerade anti-IgG sekundära antikroppar tillhandahölls av Jackson ImmunoResearch Laboratories, Inc (West Grove, PA, USA).
Försöksdjur
C57BL/6-honmöss (Harlan, Indianapolis, IN, USA) som vägde 20-25 g vid tidpunkten för försöket inhystes 5 per bur i stora skokartongburar i ett ljust (12 h ljus / mörker) och temperaturkontrollerat rum. Honmöss användes eftersom de är kända för att vara mycket känsliga för neuronala skador av de neurotoxiska amfetaminerna och för att upprätthålla konsekvens med våra tidigare studier av metamfetamins neurotoxicitet (Thomas et al. 2010, Thomas et al. 2008, Thomas et al. 2009). Mössen hade fri tillgång till mat och vatten. Institutional Care and Use Committee of Wayne State University godkände djurens vård och experimentella förfaranden. Alla förfaranden var också i överensstämmelse med NIH Guide for the Care and Use of Laboratory Animals.
Farmakologiska, fysiologiska och beteendemässiga förfaranden
Möss behandlades med mefedron med hjälp av en binge-liknande regim bestående av 4 injektioner på 10, 20 eller 40 mg/kg med ett intervall på 2 timmar mellan varje injektion. Denna binge-behandlingsregim, när den används för att injicera substituerade amfetaminer och katinonderivat, resulterar i omfattande skador på DA-nervens ändar. De doser av mefedron som används för närvarande har tidigare visat sig vara icke-toxiska för DA-nervändarna (Angoa-Perez et al. 2012). Möss behandlades med metamfetamin (4X 2,5 eller 5 mg/kg), amfetamin (4X 5 mg/kg) eller MDMA (4X 20 mg/kg) ensamt eller i kombination med mefedron. När mössen behandlades med två läkemedel fick de en mefedroninjektion 30 minuter före var och en av de fyra injektionerna av metamfetamin, amfetamin eller MDMA. Kontrollgrupperna fick injektioner av fysiologisk saltlösning enligt samma schema som användes för mefedron ensamt eller i kombination med andra amfetaminer. Som en kontroll för effekterna av en DAT-hämmare på metamfetamintoxicitet behandlades möss med nomifensin (4X 5 mg/kg) 30 minuter före varje injektion av metamfetamin (4X 5 mg/kg). Alla injektioner gavs via i.p.-vägen. Mössen avlivades 2 dagar efter den sista läkemedelsbehandlingen när den amfetaminassocierade neurotoxiciteten hade nått sitt maximum. Kroppstemperaturen övervakades genom telemetri med hjälp av IPTT-300 implanterbara temperaturtranspondrar från Bio Medic Data Systems, Inc. (Seaford, DE, USA). Temperaturerna registrerades icke-invasivt var 20:e minut med början 60 minuter före den första METH-injektionen och fortsatte därefter i 9 timmar med hjälp av DAS-5001-konsolsystemet från Bio Medic.
Bestämning av striatal DA-innehåll
Striatal vävnad dissekerades bilateralt från hjärnan efter behandling och lagrades vid -80 ° C. Frysta vävnader vägdes och sonikerades i 10 volymer 0,16 N perklorsyra vid 4 ° C. Olösligt protein avlägsnades genom centrifugering och DA bestämdes genom HPLC med elektrokemisk detektion enligt tidigare beskrivning för metamfetamin (Thomas et al. 2010, Thomas et al, 2009).
Bestämning av TH- och DAT-proteinnivåer genom immunoblotting
Effekterna av läkemedelsbehandlingar på striatala TH- och DAT-nivåer bestämdes genom immunoblotting som ett index för toxicitet för striatala DA-nervändningar. Möss offrades genom halshuggning efter behandling och striatum dissekerades bilateralt. Vävnaden förvarades vid -80°C. Fryst vävnad sönderdelades genom ultraljudsbehandling i 1% SDS vid 95°C och olösligt material sedimenterades genom centrifugering. Protein bestämdes med bicinchoninsyrametoden och lika stora mängder protein (70 μg/lane) upplöstes med SDS-polyakrylamidgelelektrofores och elektroblottades sedan på nitrocellulosa. Blottarna blockerades i Tris buffrad saltlösning innehållande Tween 20 (0,1% v/v) och 5% fettfri torrmjölk i 1 timme vid rumstemperatur. Primära antikroppar mot TH (1:1000) eller DAT (1:1000) tillsattes till blottarna och fick inkubera i 16 timmar vid 4°C. Blots tvättades 3 gånger i Tris-buffrad saltlösning för att avlägsna oreagerade antikroppar och inkuberades sedan med HRP-konjugerad anti-IgG sekundär antikropp (1:4000) under 1 timme vid rumstemperatur. Immunoreaktiva band visualiserades med förstärkt kemiluminescens och de relativa tätheterna för TH- och DAT-reaktiva band bestämdes genom avbildning med en Kodak Image Station (Carestream Molecular Systems, Rochester, NY, USA) och kvantifierades med ImageJ-programvaran (NIH).
Analys av data
Tvåvägs ANOVA utfördes för att analysera doseffekterna av metamfetamin kontra mephedrone på DA, DAT och TH. Effekterna av läkemedelsbehandlingar på striatal DA-, TH- och DAT-innehåll testades för signifikans genom envägs ANOVA följt av Tukeys multipla jämförelsetest. Resultaten av läkemedelsbehandlingar på kärnkroppstemperatur över tid analyserades med hjälp av en tvåvägs ANOVA följt av Bonferronis test för att bestämma signifikansen av skillnader i temperatur vid enskilda tider efter behandlingen. Skillnader ansågs vara signifikanta om p < 0,05. Alla statistiska analyser utfördes med hjälp av GraphPad Prism version 5.02 för Windows (GraphPad Software, San Diego, CA, USA, www.graphpad.com).
Gå till:
Resultat
Effekter av mefedron på metamfetamininducerad neurotoxicitet
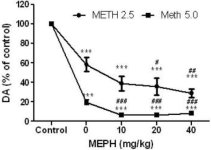
Mefedron, i doser (10, 20 eller 40 mg/kg) som man vet inte orsakar toxicitet i DA-nervens ändar (Angoa-Perez et al. 2012), administrerades 30 minuter före varje injektion av metamfetamin. Metamfetamin gavs i doser som orsakar måttlig (4X 2,5 mg/kg) eller allvarlig (4X 5 mg/kg) skada på DA-nervändarna i striatum (Thomas et al. 2004, Thomas et al. 2010). Resultaten i figur 1 visar att huvudeffekterna av metamfetamindosen (F1,40 = 66,60, p < 0,0001) och mefedrondosen (F4,40 = 131,3, p < 0,0001) på DA-nivåerna i striatum var mycket signifikanta genom tvåvägs ANOVA. Huvudeffekten av mefedron som gavs i kombination med antingen 2.5 mg / kg (F4,22 = 35.96, p <0.001) eller 5.0 mg / kg metamfetamin (F4,17 = 953.9, p <0.0001) var också mycket signifikant av envägs ANOVA. Alla behandlingar med antingen dos av metamfetamin ± mefedron orsakade signifikant större minskningar av DA jämfört med respektive kontroll (p <0.0001 för alla). Fig. 1 visar också att mefedrondoser på 20 (p < 0,01) och 40 mg/kg (p < 0,001) signifikant förstärkte de utarmande effekterna av 2,5 mg/kg metamfetamin på DA, medan alla doser av mefedron signifikant förstärkte effekterna av 5,0 mg/kg metamfetamin på DA-nivåerna (p < 0,0001 för alla).
Fig. 1
Effekter av mefedron på metamfetamininducerade minskningar av striatal DA. Möss behandlades med de angivna doserna av mefedron (MEPH) 30 minuter före varje injektion av 2.5 (-) eller 5.0 mg / kg (■) metamfetamin (METH) och offrades 2d senare för bestämning av striatala nivåer av DA med HPLC. Data är medelvärde ± SEM för 5-7 möss per grupp. Vissa felstaplar var för små för att överstiga storleken på symbolerna och visas inte synliga. ***p < 0,001 jämfört med kontroller och #p < 0,01, ##p < 0,001 eller ###p < 0,0001 jämfört med respektive dos av metamfetamin (Tukey's test för multipla jämförelser).
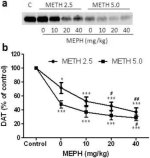
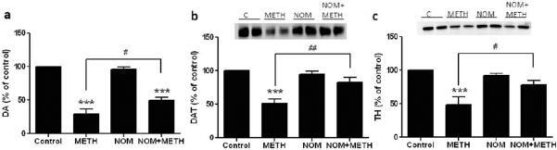
Fig. 2a visar att mefedron signifikant ökade metamfetamininducerade minskningar av DAT-nivåer som bestämts genom immunoblotting. Immunoblots kvantifierades och i överensstämmelse med resultaten för DA, huvudeffekterna av metamfetamindos (F1,92 = 9.48, p <0.001) och mefedrondos (F4,92 = 37.56, p <0.0001) på DAT-nivåer i striatum var mycket signifikanta av tvåvägs ANOVA (Fig. 2b). Huvudeffekten av mefedron som ges i kombination med antingen 2.5 mg / kg (F4,56 = 15.55, p <0.0001) eller 5.0 mg / kg metamfetamin (F4,39 = 24.84, p <0.0001) var också mycket signifikant av envägs ANOVA. Alla behandlingar med någon dos av metamfetamin ± mefedron orsakade signifikant större minskningar av DAT jämfört med respektive kontroll (p <0,01 för 2,5 mg / kg metamfetamin ensam; p <0,0001 för alla andra behandlingar). Fig. 2b visar också att mefedrondoser på 20 mg/kg (p < 0,01) och 40 mg/kg (p < 0,001) signifikant förstärkte minskningarna av DAT orsakade av 2,5 mg/kg metamfetamin, medan endast mefedrondosen på 40 mg/kg signifikant förstärkte (p < 0,01) effekterna av 5,0 mg/kg metamfetamin på minskningarna av DAT.
Fig. 2
Effekter av mefedron på metamfetamininducerade minskningar av striatal DAT. Möss behandlades med de angivna doserna av mefedron (MEPH) 30 minuter före varje injektion av 2.5 (●) eller 5.0 mg / kg (■) metamfetamin (METH) och offrades 2d senare för bestämning av striatala nivåer av DAT genom immunoblotting (a). Blots kvantifierades med hjälp av ImageJ och data är medelvärde ± SEM för 10-12 möss per grupp (b). *p <0,01 eller ***p <0,0001 vs kontroll (C) och #p <0,01 eller ##p <0,001 vs respektive dos av metamfetamin (Tukeys multipla jämförelsetest).
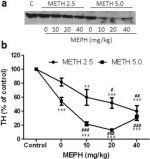
Fig. 3a visar att mefedron signifikant ökade metamfetamininducerade minskningar av TH-nivåer som bestämts genom immunoblotting. Immunoblots kvantifierades och i överensstämmelse med resultaten ovan för DA och DAT, huvudeffekterna av metamfetamindos (F1,81 = 47.89, p <0.0001) och mefedronedos (F4,81 = 63.57, p <0.0001) var mycket signifikanta av tvåvägs ANOVA (Fig. 3b). Huvudeffekten av mefedron givet i kombination med antingen 2,5 mg/kg (F4,34 = 12,98, p < 0,0001) eller 5,0 mg/kg metamfetamin (F4,49 = 99,16, p < 0,0001) var också mycket signifikant med envägs ANOVA. Alla behandlingar med endera dosen av metamfetamin ± mefedron orsakade signifikant större minskningar av TH jämfört med respektive kontroll (p < 0,001 för 2,5 mg/kg metamfetamin + 10 mg/kg mefedron; p < 0,0001 för alla andra kombinationer) med undantag för enbart 2,5 mg/kg metamfetamin som inte signifikant förändrade TH-nivåerna (dvs. ingen toxicitet). Fig. 3b visar också att mefedrondoser på 20 mg/kg (p < 0,01) och 40 mg/kg (p < 0,001) signifikant förstärkte de minskningar av TH som orsakades av 2,5 mg/kg metamfetamin och att alla tre mefedrondoserna signifikant (p < 0,0001) förstärkte effekterna av 5,0 mg/kg metamfetamin på TH-minskningarna.
Fig. 3: Effekter av mefedron på metamfetamin
Effekter av mefedron på metamfetamininducerade minskningar av striatal TH. Möss behandlades med de angivna doserna av mefedron (MEPH) 30 minuter före varje injektion av 2,5 (●) eller 5,0 mg/kg (■) metamfetamin (METH) och offrades 2d senare för bestämning av striatala nivåer av TH genom immunoblotting (a). Blots kvantifierades med hjälp av ImageJ och data är medelvärde ± SEM för 10-12 möss per grupp (b). Vissa felstaplar var för små för att överstiga storleken på symbolerna och visas inte synliga. **p <0,001 eller ***p <0,0001 vs kontroll (C) och #p <0,01, ##p <0,001 eller ###p <0,0001) vs respektive dos av metamfetamin (Tukeys multipla jämförelsetest).
Effekter av mefedron på metamfetamininducerad hypertermi
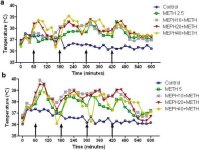
Mefedron, liksom metamfetamin, orsakar betydande hypertermi (Hadlock et al. 2011, Baumann et al. 2012, Angoa-Perez et al. 2012). När mefedron gavs 30 minuter före varje injektion av metamfetamin, kan man i figur 4 se att huvudeffekterna av metamfetamin- och mefedrondoser (F1,300 = 11,99, p < 0,0001) över tid (F4,300 = 51,73, p < 0,0001) var mycket signifikanta genom tvåvägs ANOVA. Huvudeffekterna av mefedron givet i kombination med antingen 2,5 mg/kg metamfetamin (F4,120 = 41,44, p < 0,0001, panel a) över tid (F30,120 = 3,84, p < 0,0001) eller 5,0 mg/kg metamfetamin (F4,120 = 78,09, p < 0,0001, panel b) över tid (F30,120 = 9,98, p < 0,0001) var också mycket signifikanta genom tvåvägs ANOVA. Alla behandlingar med någon dos av metamfetamin ± mefedron skilde sig signifikant från respektive kontroller (p <0,0001 för alla behandlingar).
Fig. 4
Effekter av mefedron på metamfetamininducerad hypertermi. Möss behandlades med de angivna doserna av mefedron (MEPH) 30 minuter före varje injektion av 2,5 (a) eller 5,0 mg/kg (b) metamfetamin (METH). Kärntemperaturen mättes med 20 minuters intervall med telemetri med början 60 minuter före den första injektionen av metamfetamin. De 4 metamfetamininjektionerna indikeras av pilarna som vilar på x-axeln. Data uttrycks som genomsnittlig kroppstemperatur för 6-8 möss per grupp. SEM var alltid <10% av medelvärdet och utelämnas för tydlighetens skull.
Effekter av mefedron på amfetamin- och MDMA-inducerad neurotoxicitet
För att testa om de förstärkande effekterna av mefedron på metamfetamin kunde utsträckas till andra neurotoxiska amfetaminer, behandlades möss med denna β-ketoamfetamin (20 mg/kg) plus amfetamin (4X 5 mg/kg) eller MDMA (4X 20 mg/kg) och resultaten presenteras i figur 5. Kom ihåg att mefedron i sig inte minskar striatal DA, DAT eller TH (Angoa-Perez et al. 2012). Huvudeffekten av läkemedel (F5,27 = 27.18, p <0.0001) var mycket signifikant genom envägs ANOVA för DA-reduktioner (Fig. 5a). Det kan också ses i Fig. 5a att alla behandlingar med amfetamin (p <0.001) eller MDMA (p <0.001) ensam eller i kombination med mephedrone (p <0.0001 för båda läkemedlen) sänkte DA-nivåerna signifikant från kontroll. Mephedrone förbättrade signifikant DA-reduktioner orsakade av amfetamin (p <0.01) eller MDMA (p <0.01). Fig. 5b visar liknande effekter av kombinationsläkemedelsbehandlingar på DAT-nivåer i striatum. Huvudeffekten av läkemedel (F4,49 = 42.63, p <0.0001) var mycket signifikant av envägs ANOVA för DAT. Det kan också ses i fig. 5b att alla behandlingar med amfetamin eller MDMA var signifikant (p <0.0001 för alla) lägre jämfört med kontroll. Mephedrone förbättrade också signifikant DAT-reduktioner orsakade av antingen amfetamin eller MDMA (p <0.0001 i båda fallen). Slutligen, Fig. 5c visar att läkemedlets huvudeffekt (F4,50 = 75.06, p <0.0001) var mycket signifikant av envägs ANOVA för minskningar i TH. Det kan också ses i fig. 5c att alla behandlingar med amfetamin eller MDMA var signifikant (p < 0,0001 för alla) lägre i jämförelse med kontroll. Mephedrone förbättrade också signifikant TH-reduktioner orsakade av antingen amfetamin eller MDMA (p <0.0001 i båda fallen)
Fig. 5
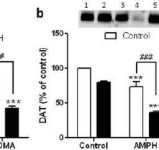
Effekter av mefedron på amfetamin- eller MDMA-inducerad DA nervändande neurotoxicitet. Möss behandlades med 20 mg/kg mefedron (MEPH) 30 minuter före varje injektion av 5,0 mg/kg amfetamin (AMPH) eller 20 mg/kg MDMA och offrades 2d efter behandling för bestämning av striatala nivåer av (a) DA med HPLC. (b) DAT och (c) TH bestämdes genom immunoblotting och blottar kvantifierades med hjälp av ImageJ. Representativa immunoblots för DAT och TH ingår som insatser i paneler (b) respektive (c) och behandlingar för båda panelerna indikeras med 1,5: kontroll; 2,6: MEPH; 3: AMPH; 4: AMPH + MEPH; 7: MDMA; och 8: MDMA + MEPH. Data är medelvärde ± SEM för 5-12 möss i varje grupp. **p <0,001 eller ***p <0,0001 vs kontroll och #p <0,01 eller ###p <0,0001 vs AMPH eller MDMA (Tukey's multipla jämförelsetest).
Effekter av nomifensin på metamfetamininducerad neurotoxicitet
Nomifensine, en kraftfull DAT-blockerare utan känd missbruks- eller neurotoxisk potential, testades för sin förmåga att skydda mot metamfetamininducerad neurotoxicitet och för kontrast till mefedronets verkan på toxiciteten för DA-nervändningar orsakade av metamfetamin, amfetamin och MDMA. Resultaten i fig. 6a visar att läkemedlets huvudeffekt (F3,16 = 63,39, p < 0,0001) på DA-nivåer var mycket signifikant genom envägs ANOVA. Nomifensine ensam förändrade inte DA-nivåerna men minskningen orsakad av metamfetamin (p <0,0001) reverserades något men signifikant av nomifensine (p <0,01). Huvudeffekten av läkemedel (F3,20 = 16.78, p <0.0001) på DAT-nivåer var mycket signifikant av envägs ANOVA som visas i Fig. 6b. Nomifensine förändrade inte DAT-nivåer men gav signifikant skydd (p <0.001) mot minskningen av striatal DAT orsakad av metamfetamin (p <0.0001) jämfört med kontroll. Slutligen, Fig. 6c visar att läkemedlets huvudeffekt (F3,15 = 14.10, p <0.0001) på TH-nivåer var mycket signifikant av envägs ANOVA. Som sett för DA och DAT, minskningen av TH orsakad av metamfetamin (p <0.0001) förhindrades något men signifikant av nomifensin (p <0.01).
Fig. 6
Effekter av nomifensin på metamfetamininducerad DA nervändande neurotoxicitet. Möss behandlades med 5,0 mg/kg nomifensin (NOM) 30 minuter före varje injektion av 5,0 mg/kg metamfetamin (METH) och offrades 2 dagar senare för bestämning av striatala nivåer av (a) DA med HPLC. (b) DAT och (c) TH bestämdes genom immunoblotting och blottar kvantifierades med hjälp av ImageJ. Representativa immunoblots för DAT och TH ingår som insatser i panelerna (b) och (c) respektive. Data är medelvärde plus SEM för 5-7 möss per grupp. ***p <0,0001 vs kontroll (C) och #p <0,01 eller ##p <0,001 vs enbart metamfetamin (Tukeys multipla jämförelsetest).
Diskussion
Syftet med den aktuella studien var att avgöra om mefedron skulle förhindra DA-nervavslutande toxicitet orsakad av metamfetamin. Baserat på dess kemiska likhet med metamfetamin och MDMA förväntades det ursprungligen att mefedron skulle utöva skadliga effekter på DA-neuronsystemet. Flera studier visade dock nästan samtidigt att mefedron inte var giftigt för DA-nervändarna (Angoa-Perez et al. 2012, Baumann et al. 2012, Hadlock et al. 2011). Frågan om huruvida detta läkemedel orsakar skador på 5-HT-neuronsystemet förblir öppen. En studie rapporterade ihållande minskningar av 5-HT-nervändfunktion (Hadlock et al. 2011) medan en annan fann att mefedron inte orsakade skada (Baumann et al. 2012). Mefedron interagerar med DA-nervänden på ett sätt som tyder på att det verkligen stimulerar frisättning och blockerar DA-återupptag via dess interaktioner med DAT. En viktig aspekt av metamfetamins neurotoxiska verkningsmekanism är dess förmåga att få tillgång till DA-nervändarna genom DAT och störa DA-homeostasen (Sulzer 2011). Om detta tidiga steg i metamfetaminets neurotoxiska kaskad förhindras genom hämning av DAT, förhindras toxicitet (Pu et al. 1994, Poth et al. 2012, Marek et al. 1990, Schmidt och Gibb 1985). Vi antog att mefedron skulle kunna ha samma skyddande egenskaper som andra DAT-hämmare, men observerade istället en signifikant ökning av toxiciteten. Denna interaktion sågs med hjälp av två olika doser av metamfetamin som orsakar måttlig eller allvarlig skada på DA-nervändarna (4X 2,5 respektive 5,0 mg/kg). Denna förstärkande effekt av mefedron var inte begränsad till metamfetamin utan utsträcktes till amfetamin och MDMA, två droger som ofta sammissbrukas med mefedron och andra β-ketoamfetaminer (Feyissa och Kelly 2008, Schifano et al. 2011, Kelly 2011). Trots att mefedron inte orsakar toxicitet för åtminstone DA-nervändarna i striatum förstärker det därför de neurotoxiska effekterna av andra missbrukssubstanser. Detta nya fynd bör ge mefedronmissbruk en ännu skarpare bild, eftersom dess brist på inneboende neurotoxicitet kan få det att framstå som ofarligt.
Hypertermi är en ofta rapporterad akut biverkning av intag av metamfetamin (Greene et al. 2008) och β-ketoamfetamin hos människor (Borek och Holstege 2012, Prosser och Nelson 2012). Liksom metamfetamin orsakar många av β-ketoamfetaminläkemedlen också betydande höjningar av kärntemperaturen hos gnagare (Angoa-Perez m.fl. 2012, Hadlock m.fl. 2011, Baumann m.fl. 2012, Rockhold m.fl. 1997). Även om den hypertermi som orsakas av metamfetamin kan bidra till dess morfologiska och neuronala skadeverkningar, är det inte nödvändigtvis så att hypertermi är den direkta orsaken till dessa effekter (Kiyatkin och Sharma 2009). Vi registrerade kroppstemperaturen hos möss som behandlats med mefedron och metamfetamin och observerade att den kombinerade behandlingen inte ökade temperaturen utöver de maximala ökningar som sågs efter endera läkemedlet ensamt. Metamfetamin orsakade en dosrelaterad ökning av kroppstemperaturen och denna hypertermi var oföränderlig över hela det testade dosintervallet för mefedron. Faktum är att det fall i kroppstemperatur efter injektion som observerades efter mefedronbehandling (Angoa-Perez et al. 2012) bibehölls vid högre doser av mefedron plus metamfetamin. Även om den läkemedelsinducerade hypertermi inte förstärktes av kombinerad läkemedelsbehandling, var de neurotoxiska effekterna additiva. Därför förefaller det, åtminstone i det aktuella fallet, som om de neurotoxiska effekterna av metamfetamin kan förstärkas av mefedron på ett sätt som är oberoende av hypertermi.
Mefedron hämmar tydligt DAT-funktionen och blockerar återupptaget av DA in vitro (Lopez-Arnau et al. 2012, Hadlock et al. 2011, Kehr et al. 2011, Martinez-Clemente et al. 2012, Cozzi et al. 1999). Mefedron undantränger WIN-35 428 från dess bindningsställe på DAT, vilket tyder på att det är en konkurrenskraftig hämmare av DA-upptag (Martinez-Clemente et al. 2012, Lopez-Arnau et al. 2012). Mefedronets styrka i detta avseende är mycket lik metamfetaminets (Cozzi et al. 1999) och MDMA:s (Escubedo et al. 2011). Det är inte känt om mefedron transporteras av DAT, men det gör metkatinon (Cozzi och Foley 2003). Nomifensin och amfonsyra, som binder till DAT och hämmar DA-upptaget, ger ett betydande skydd mot metamfetamininducerad neurotoxicitet (Pu et al. 1994, Marek et al. 1990, Schmidt och Gibb 1985, Poth et al. 2012) och möss som saknar DAT är resistenta mot metamfetaminets neuronala toxicitet (Fumagalli et al. 1998). Att veta att mefedron är icke-neurotoxiskt och en DAT-blockerare leder till förutsägelsen att det borde förhindra toxicitet. Vi testade nomifensine i detta avseende som en positiv kontroll och bekräftade att det skyddar mot metamfetamininducerad utarmning av DA, DAT och TH. Nomifensine hämmar också norepinefrintransportören (Brogden et al. 1979) men denna egenskap kan inte förklara de nuvarande resultaten eftersom de flesta β-ketoamfetaminer inklusive mefedron hämmar norepinefrintransportören och blockerar norepinefrinupptag (Kelly 2011, Rothman et al. 2003, Cozzi et al. 1999, Sogawa et al. 2011, Lopez-Arnau et al. 2012). En roll för det neuronala 5-HT-systemet i vissa av mefedronets farmakologiska verkningar är möjlig mot bakgrund av detta läkemedels förmåga att, liksom MDMA (Yamamoto et al. 1995), orsaka utflöde av striatal DA via dess interaktioner med 5-HT2A-receptorer (Lopez-Arnau et al. 2012, Martinez-Clemente et al. 2012). Den hyperlokomotion som orsakas av mefedron är beroende av endogent 5-HT (Lopez-Arnau et al. 2012) och detta läkemedel stimulerar också frisättningen av 5-HT och hämmar dess upptag in vitro (Sogawa et al. 2011, Cozzi et al. 1999, Nagai et al. 2007, Hadlock et al. 2011, Lopez-Arnau et al. 2012, Martinez-Clemente et al. 2012) och in vivo (Baumann et al. 2012, Kehr et al. 2011). Vi kan dock utesluta en roll för endogent 5-HT i DA-neurotoxiciteten hos åtminstone metamfetamin genom att visa att möss som är genetiskt utarmade på 5-HT behåller sin känslighet för neurotoxicitet (Thomas et al. 2010).
Mefedron kan förstärka metamfetamins neurotoxicitet genom flera möjliga mekanismer. För det första kan mefedron interagera med VMAT för att orsaka läckage av DA till cytoplasman i den presynaptiska nervändan. Behandlingar som ökar den cytoplasmatiska poolen (dvs. läkemedelsfrisättande) av DA ökar metamfetaminets neurotoxicitet (Thomas et al. 2008, Thomas et al. 2009, Schmidt et al. 1985). Denna mekanism är inte sannolik eftersom metkatinon endast interagerar svagt med VMAT (Cozzi et al. 1999). För det andra kan kombinationen av mefedron plus metamfetamin ha en synergistisk effekt på icke-vesikulär frisättning av DA, men denna möjlighet verkar också osannolik mot bakgrund av resultat som visar att behandling av DAT- eller SERT-uttryckande CHO-celler med metylon plus metamfetamin inte har en additiv effekt på DA- eller 5-HT-frisättning (Sogawa et al. 2011). För det tredje kan mefedron interagera med DAT på ett nytt sätt som bidrar till additiv toxicitet. Det har visats att metylon i kombination med metamfetamin orsakar synergistisk cytotoxicitet i CHO-celler som uttrycker DAT eller SERT men inte i CHO-celler av vildtyp som saknar transportörer (Sogawa et al. 2011). Den cytotoxicitet som ses i odlade celler i dessa studier (dvs. LDH-frisättning) skiljer sig mycket från skadorna på DA-nervändarna som orsakas av metamfetamin, men denna mekanism antyder en intressant men odefinierad roll för DAT i ökad cytotoxicitet. Slutligen kan mefedron förändra metamfetaminets metabolism. Mefedron metaboliseras främst genom N-demetylering (Meyer och Maurer 2010), liksom metamfetamin och MDMA (Caldwell 1976). Stöd för denna mekanism kommer från demonstrationen att metamfetamin och MDMA ömsesidigt hämmar produktionen av sina respektive primära metaboliter och höjer plasmanivåerna av läkemedlen över dem som ses efter administrering av endera läkemedlet ensamt (Kuwayama et al. 2012). De doser av mefedron som används för närvarande och i vår tidigare studie (Angoa-Perez et al. 2012) är visserligen höga men inte neurotoxiska och ligger inom det intervall som missbrukas av människor (McErath och O'Neill 2011). Därför kan mefedron fungera som MDMA för att öka plasmanivåerna av metamfetamin genom att hämma dess metabolism. En fördjupad farmakokinetisk analys kommer att krävas för att bekräfta denna senare möjlighet.
Missbruket av β-ketoamfetaminer ökar i en alarmerande takt och mefedron är nu en av de vanligaste drogerna efter cannabis, MDMA och kokain (Morris 2010, Winstock et al. 2011b). Dessutom framkallar mefedron starkare känslor av begär hos människor jämfört med MDMA (Brunt m.fl. 2011) och användare som snortar mefedron anser att det är mer beroendeframkallande än kokain (Winstock m.fl. 2011b). Mefedron konsumeras av människor på ett binge-liknande sätt (dvs. "stacking") och tas ofta tillsammans med andra droger som cannabis och amfetaminpsykostimulantia (Schifano m.fl. 2011, Fass m.fl. 2012, Winstock m.fl. 2011a, Kelly 2011, Torrance och Cooper 2010). Mefedron förekommer allt oftare i tabletter som säljs som MDMA (Brunt m.fl. 2011) och användningen av mefedron kommer sannolikt att överstiga användningen av MDMA i takt med att renheten hos den senare drogen fortsätter att sjunka (Brunt m.fl. 2011, Tanner-Smith 2006, Teng m.fl. 2006). Baserat på de vanliga missbruksmönstren för mefedron och andra ingredienser i "badsalt" är det viktigt att överväga om ytterligare hälsorisker uppstår hos människor när dessa droger avsiktligt eller oavsiktligt kombineras med amfetamin. Våra resultat som visar att åtminstone mefedron avsevärt förstärker neurotoxiciteten för DA-nervändarna i striatum som orsakas av metamfetamin, amfetamin och MDMA avslöjar en särskilt farlig och oväntad egenskap hos denna β-ketoamfetamin.
Förkortningar som används
5-HT serotonin
DA dopamin
DAT DA-transportör
MDMA 3,4-metylendioximetamfetamin
TH tyrosinhydroxylas
VMAT vesikulär monoamintransportör
Mefedron (4-metylmetkatinon) är ett katinonderivat och en strukturell analog till metamfetamin och 3,4-metylendioximetamfetamin (MDMA). Mefedron är en psykoaktiv ingrediens i "badsalter" tillsammans med andra föreningar som metylon, butylon och 3,4-metylendioxypyrovaleron (MDPV). β-ketoamfetaminer missbrukas i allt högre grad, vilket till stor del beror på den mycket begränsade tillgången på prekursorer som behövs för syntes av metamfetamin och MDMA i hemliga laboratorier och en motsvarande minskning av deras renhet (Winstock m.fl. 2011b, Brunt m.fl. 2011). I takt med att missbruket av β-ketoamfetaminer fortsätter att öka har listan över deras negativa effekter utökats till att omfatta kardiovaskulära komplikationer, agitation, sömnlöshet, psykoser och depression (Schifano m.fl. 2011, Prosser och Nelson 2012).
Som kemiska släktingar till metamfetamin och MDMA är det inte förvånande att β-ketoamfetaminerna har många av samma effekter på det centrala nervsystemet som de förstnämnda drogerna. Till exempel blockerar dessa läkemedel dopamin- (DA) och serotonin- (5-HT) transportörer (DAT respektive SERT) (Cozzi et al. 1999, Rothman et al. 2003, Fleckenstein et al. 2000, Lopez-Arnau et al. 2012) och de stimulerar monoaminfrisättning in vitro (Kalix och Glennon 1986, Gygi et al. 1997, Rothman et al. 2003) och in vivo (Gygi et al. 1997, Kehr et al. 2011). Metkatinon orsakar ihållande minskningar av tryptofanhydroxylas- och tyrosinhydroxylas (TH)-aktivitet och utarmning av DA och 5-HT (Gygi et al. 1997, Gygi et al. 1996, Sparago et al. 1996). PET-avbildningsstudier hos avhållsamma metkatinonanvändare avslöjade minskad DAT-densitet i striatal, vilket tyder på en förlust av DA-terminaler (McCann et al. 1998). Den samtidiga stimuleringen av DA-frisättning och hämningen av dess upptag speglar de kritiska element som ligger bakom neurotoxiciteten i samband med metamfetamin (Kuhn et al. 2008, Yamamoto och Bankson 2005, Cadet et al. 2007, Fleckenstein et al. 2007).
Vi (Angoa-Perez et al. 2012) och andra (Baumann et al. 2012, Hadlock et al. 2011) har nyligen undersökt möjligheten att mefedron skulle kunna orsaka neurotoxicitet på samma sätt som metamfetamin och MDMA. Överraskande nog var mefedron inte giftigt för DA-nervändarna i striatum (Hadlock et al. 2011, Baumann et al. 2012, Angoa-Perez et al. 2012). Frågan om huruvida mefedron skadar 5-HT-nervändarna är fortfarande oklar eftersom en studie dokumenterade positiva effekter (Hadlock et al. 2011) medan en annan var negativ (Baumann et al. 2012). Mot bakgrund av den relativt godartade effekten av mefedron på DA-nervändar och med tanke på dess egenskaper som DAT-blockerare, antog vi att det faktiskt skulle kunna skydda DA-neuronsystemet från de neurotoxiska effekterna av metamfetamin, precis som det är känt att det förekommer med andra DAT-blockerare såsom amfonsyra (Pu et al. 1994, Schmidt och Gibb 1985, Marek et al. 1990) och nomifensin (Poth et al. 2012). Vi rapporterar för närvarande att mefedron avsevärt förstärker neurotoxiciteten hos metamfetamin. Denna effekt gäller även amfetamin och MDMA, droger som ofta missbrukas tillsammans med mefedron (Feyissa och Kelly 2008, Schifano et al. 2011). Dessa överraskande resultat sätter missbruket av mefedron i ett nytt ljus och gör det angeläget att uppmärksamma denna subtila och farliga egenskap hos denna β-ketoamfetamin.
Material och metoder
Läkemedel och reagenser
Mefedronhydroklorid och 3,4-metylendioxymetamfetamin (MDMA) erhölls från NIDA Research Resources Drug Supply Program. (+) Metamfetaminhydroklorid, nomifensinmaleat, d-amfetaminsulfat, pentobarbital, DA och alla buffertar och HPLC-reagenser köptes från Sigma-Aldrich (St. Louis, MO, USA). Bicinchoninic acid protein assay kit erhölls från Pierce (Rockford, IL, USA). Polyklonala antikroppar mot TH från råttor framställdes enligt tidigare beskrivning (Kuhn och Billingsley 1987). Monoklonala antikroppar mot DAT från råtta tillhandahölls generöst av Dr. Roxanne A. Vaughan (University of North Dakota, Grand Forks, ND, USA). HRP-konjugerade anti-IgG sekundära antikroppar tillhandahölls av Jackson ImmunoResearch Laboratories, Inc (West Grove, PA, USA).
Försöksdjur
C57BL/6-honmöss (Harlan, Indianapolis, IN, USA) som vägde 20-25 g vid tidpunkten för försöket inhystes 5 per bur i stora skokartongburar i ett ljust (12 h ljus / mörker) och temperaturkontrollerat rum. Honmöss användes eftersom de är kända för att vara mycket känsliga för neuronala skador av de neurotoxiska amfetaminerna och för att upprätthålla konsekvens med våra tidigare studier av metamfetamins neurotoxicitet (Thomas et al. 2010, Thomas et al. 2008, Thomas et al. 2009). Mössen hade fri tillgång till mat och vatten. Institutional Care and Use Committee of Wayne State University godkände djurens vård och experimentella förfaranden. Alla förfaranden var också i överensstämmelse med NIH Guide for the Care and Use of Laboratory Animals.
Farmakologiska, fysiologiska och beteendemässiga förfaranden
Möss behandlades med mefedron med hjälp av en binge-liknande regim bestående av 4 injektioner på 10, 20 eller 40 mg/kg med ett intervall på 2 timmar mellan varje injektion. Denna binge-behandlingsregim, när den används för att injicera substituerade amfetaminer och katinonderivat, resulterar i omfattande skador på DA-nervens ändar. De doser av mefedron som används för närvarande har tidigare visat sig vara icke-toxiska för DA-nervändarna (Angoa-Perez et al. 2012). Möss behandlades med metamfetamin (4X 2,5 eller 5 mg/kg), amfetamin (4X 5 mg/kg) eller MDMA (4X 20 mg/kg) ensamt eller i kombination med mefedron. När mössen behandlades med två läkemedel fick de en mefedroninjektion 30 minuter före var och en av de fyra injektionerna av metamfetamin, amfetamin eller MDMA. Kontrollgrupperna fick injektioner av fysiologisk saltlösning enligt samma schema som användes för mefedron ensamt eller i kombination med andra amfetaminer. Som en kontroll för effekterna av en DAT-hämmare på metamfetamintoxicitet behandlades möss med nomifensin (4X 5 mg/kg) 30 minuter före varje injektion av metamfetamin (4X 5 mg/kg). Alla injektioner gavs via i.p.-vägen. Mössen avlivades 2 dagar efter den sista läkemedelsbehandlingen när den amfetaminassocierade neurotoxiciteten hade nått sitt maximum. Kroppstemperaturen övervakades genom telemetri med hjälp av IPTT-300 implanterbara temperaturtranspondrar från Bio Medic Data Systems, Inc. (Seaford, DE, USA). Temperaturerna registrerades icke-invasivt var 20:e minut med början 60 minuter före den första METH-injektionen och fortsatte därefter i 9 timmar med hjälp av DAS-5001-konsolsystemet från Bio Medic.
Bestämning av striatal DA-innehåll
Striatal vävnad dissekerades bilateralt från hjärnan efter behandling och lagrades vid -80 ° C. Frysta vävnader vägdes och sonikerades i 10 volymer 0,16 N perklorsyra vid 4 ° C. Olösligt protein avlägsnades genom centrifugering och DA bestämdes genom HPLC med elektrokemisk detektion enligt tidigare beskrivning för metamfetamin (Thomas et al. 2010, Thomas et al, 2009).
Bestämning av TH- och DAT-proteinnivåer genom immunoblotting
Effekterna av läkemedelsbehandlingar på striatala TH- och DAT-nivåer bestämdes genom immunoblotting som ett index för toxicitet för striatala DA-nervändningar. Möss offrades genom halshuggning efter behandling och striatum dissekerades bilateralt. Vävnaden förvarades vid -80°C. Fryst vävnad sönderdelades genom ultraljudsbehandling i 1% SDS vid 95°C och olösligt material sedimenterades genom centrifugering. Protein bestämdes med bicinchoninsyrametoden och lika stora mängder protein (70 μg/lane) upplöstes med SDS-polyakrylamidgelelektrofores och elektroblottades sedan på nitrocellulosa. Blottarna blockerades i Tris buffrad saltlösning innehållande Tween 20 (0,1% v/v) och 5% fettfri torrmjölk i 1 timme vid rumstemperatur. Primära antikroppar mot TH (1:1000) eller DAT (1:1000) tillsattes till blottarna och fick inkubera i 16 timmar vid 4°C. Blots tvättades 3 gånger i Tris-buffrad saltlösning för att avlägsna oreagerade antikroppar och inkuberades sedan med HRP-konjugerad anti-IgG sekundär antikropp (1:4000) under 1 timme vid rumstemperatur. Immunoreaktiva band visualiserades med förstärkt kemiluminescens och de relativa tätheterna för TH- och DAT-reaktiva band bestämdes genom avbildning med en Kodak Image Station (Carestream Molecular Systems, Rochester, NY, USA) och kvantifierades med ImageJ-programvaran (NIH).
Analys av data
Tvåvägs ANOVA utfördes för att analysera doseffekterna av metamfetamin kontra mephedrone på DA, DAT och TH. Effekterna av läkemedelsbehandlingar på striatal DA-, TH- och DAT-innehåll testades för signifikans genom envägs ANOVA följt av Tukeys multipla jämförelsetest. Resultaten av läkemedelsbehandlingar på kärnkroppstemperatur över tid analyserades med hjälp av en tvåvägs ANOVA följt av Bonferronis test för att bestämma signifikansen av skillnader i temperatur vid enskilda tider efter behandlingen. Skillnader ansågs vara signifikanta om p < 0,05. Alla statistiska analyser utfördes med hjälp av GraphPad Prism version 5.02 för Windows (GraphPad Software, San Diego, CA, USA, www.graphpad.com).
Gå till:
Resultat
Effekter av mefedron på metamfetamininducerad neurotoxicitet
Mefedron, i doser (10, 20 eller 40 mg/kg) som man vet inte orsakar toxicitet i DA-nervens ändar (Angoa-Perez et al. 2012), administrerades 30 minuter före varje injektion av metamfetamin. Metamfetamin gavs i doser som orsakar måttlig (4X 2,5 mg/kg) eller allvarlig (4X 5 mg/kg) skada på DA-nervändarna i striatum (Thomas et al. 2004, Thomas et al. 2010). Resultaten i figur 1 visar att huvudeffekterna av metamfetamindosen (F1,40 = 66,60, p < 0,0001) och mefedrondosen (F4,40 = 131,3, p < 0,0001) på DA-nivåerna i striatum var mycket signifikanta genom tvåvägs ANOVA. Huvudeffekten av mefedron som gavs i kombination med antingen 2.5 mg / kg (F4,22 = 35.96, p <0.001) eller 5.0 mg / kg metamfetamin (F4,17 = 953.9, p <0.0001) var också mycket signifikant av envägs ANOVA. Alla behandlingar med antingen dos av metamfetamin ± mefedron orsakade signifikant större minskningar av DA jämfört med respektive kontroll (p <0.0001 för alla). Fig. 1 visar också att mefedrondoser på 20 (p < 0,01) och 40 mg/kg (p < 0,001) signifikant förstärkte de utarmande effekterna av 2,5 mg/kg metamfetamin på DA, medan alla doser av mefedron signifikant förstärkte effekterna av 5,0 mg/kg metamfetamin på DA-nivåerna (p < 0,0001 för alla).
http://bbzzzsvqcrqtki6umym6itiixfhni37ybtt7mkbjyxn2pgllzxf2qgyd.onion/index.php?attachments/sywobkqrnd-jpg.47/&hash=f31155ada4e59f22a35f9110aaa6a7e5
Fig. 1
Effekter av mefedron på metamfetamininducerade minskningar av striatal DA. Möss behandlades med de angivna doserna av mefedron (MEPH) 30 minuter före varje injektion av 2.5 (-) eller 5.0 mg / kg (■) metamfetamin (METH) och offrades 2d senare för bestämning av striatala nivåer av DA med HPLC. Data är medelvärde ± SEM för 5-7 möss per grupp. Vissa felstaplar var för små för att överstiga storleken på symbolerna och visas inte synliga. ***p < 0,001 jämfört med kontroller och #p < 0,01, ##p < 0,001 eller ###p < 0,0001 jämfört med respektive dos av metamfetamin (Tukey's test för multipla jämförelser).
Fig. 2a visar att mefedron signifikant ökade metamfetamininducerade minskningar av DAT-nivåer som bestämts genom immunoblotting. Immunoblots kvantifierades och i överensstämmelse med resultaten för DA, huvudeffekterna av metamfetamindos (F1,92 = 9.48, p <0.001) och mefedrondos (F4,92 = 37.56, p <0.0001) på DAT-nivåer i striatum var mycket signifikanta av tvåvägs ANOVA (Fig. 2b). Huvudeffekten av mefedron som ges i kombination med antingen 2.5 mg / kg (F4,56 = 15.55, p <0.0001) eller 5.0 mg / kg metamfetamin (F4,39 = 24.84, p <0.0001) var också mycket signifikant av envägs ANOVA. Alla behandlingar med någon dos av metamfetamin ± mefedron orsakade signifikant större minskningar av DAT jämfört med respektive kontroll (p <0,01 för 2,5 mg / kg metamfetamin ensam; p <0,0001 för alla andra behandlingar). Fig. 2b visar också att mefedrondoser på 20 mg/kg (p < 0,01) och 40 mg/kg (p < 0,001) signifikant förstärkte minskningarna av DAT orsakade av 2,5 mg/kg metamfetamin, medan endast mefedrondosen på 40 mg/kg signifikant förstärkte (p < 0,01) effekterna av 5,0 mg/kg metamfetamin på minskningarna av DAT.
http://bbzzzsvqcrqtki6umym6itiixfhni37ybtt7mkbjyxn2pgllzxf2qgyd.onion/index.php?attachments/bfu3sx5dnr-jpg.4797/&hash=f31155ada4e59f22a35f9110aaa6a7e5
Fig. 2
Effekter av mefedron på metamfetamininducerade minskningar av striatal DAT. Möss behandlades med de angivna doserna av mefedron (MEPH) 30 minuter före varje injektion av 2.5 (●) eller 5.0 mg / kg (■) metamfetamin (METH) och offrades 2d senare för bestämning av striatala nivåer av DAT genom immunoblotting (a). Blots kvantifierades med hjälp av ImageJ och data är medelvärde ± SEM för 10-12 möss per grupp (b). *p <0,01 eller ***p <0,0001 vs kontroll (C) och #p <0,01 eller ##p <0,001 vs respektive dos av metamfetamin (Tukeys multipla jämförelsetest).
Fig. 3a visar att mefedron signifikant ökade metamfetamininducerade minskningar av TH-nivåer som bestämts genom immunoblotting. Immunoblots kvantifierades och i överensstämmelse med resultaten ovan för DA och DAT, huvudeffekterna av metamfetamindos (F1,81 = 47.89, p <0.0001) och mefedronedos (F4,81 = 63.57, p <0.0001) var mycket signifikanta av tvåvägs ANOVA (Fig. 3b). Huvudeffekten av mefedron givet i kombination med antingen 2,5 mg/kg (F4,34 = 12,98, p < 0,0001) eller 5,0 mg/kg metamfetamin (F4,49 = 99,16, p < 0,0001) var också mycket signifikant med envägs ANOVA. Alla behandlingar med endera dosen av metamfetamin ± mefedron orsakade signifikant större minskningar av TH jämfört med respektive kontroll (p < 0,001 för 2,5 mg/kg metamfetamin + 10 mg/kg mefedron; p < 0,0001 för alla andra kombinationer) med undantag för enbart 2,5 mg/kg metamfetamin som inte signifikant förändrade TH-nivåerna (dvs. ingen toxicitet). Fig. 3b visar också att mefedrondoser på 20 mg/kg (p < 0,01) och 40 mg/kg (p < 0,001) signifikant förstärkte de minskningar av TH som orsakades av 2,5 mg/kg metamfetamin och att alla tre mefedrondoserna signifikant (p < 0,0001) förstärkte effekterna av 5,0 mg/kg metamfetamin på TH-minskningarna.
http://bbzzzsvqcrqtki6umym6itiixfhni37ybtt7mkbjyxn2pgllzxf2qgyd.onion/index.php?attachments/tggfrjpqru-jpg.4798/&hash=f31155ada4e59f22a35f9110aaa6a7e5
Fig. 3: Effekter av mefedron på metamfetamin
Effekter av mefedron på metamfetamininducerade minskningar av striatal TH. Möss behandlades med de angivna doserna av mefedron (MEPH) 30 minuter före varje injektion av 2,5 (●) eller 5,0 mg/kg (■) metamfetamin (METH) och offrades 2d senare för bestämning av striatala nivåer av TH genom immunoblotting (a). Blots kvantifierades med hjälp av ImageJ och data är medelvärde ± SEM för 10-12 möss per grupp (b). Vissa felstaplar var för små för att överstiga storleken på symbolerna och visas inte synliga. **p <0,001 eller ***p <0,0001 vs kontroll (C) och #p <0,01, ##p <0,001 eller ###p <0,0001) vs respektive dos av metamfetamin (Tukeys multipla jämförelsetest).
Effekter av mefedron på metamfetamininducerad hypertermi
Mefedron, liksom metamfetamin, orsakar betydande hypertermi (Hadlock et al. 2011, Baumann et al. 2012, Angoa-Perez et al. 2012). När mefedron gavs 30 minuter före varje injektion av metamfetamin, kan man i figur 4 se att huvudeffekterna av metamfetamin- och mefedrondoser (F1,300 = 11,99, p < 0,0001) över tid (F4,300 = 51,73, p < 0,0001) var mycket signifikanta genom tvåvägs ANOVA. Huvudeffekterna av mefedron givet i kombination med antingen 2,5 mg/kg metamfetamin (F4,120 = 41,44, p < 0,0001, panel a) över tid (F30,120 = 3,84, p < 0,0001) eller 5,0 mg/kg metamfetamin (F4,120 = 78,09, p < 0,0001, panel b) över tid (F30,120 = 9,98, p < 0,0001) var också mycket signifikanta genom tvåvägs ANOVA. Alla behandlingar med någon dos av metamfetamin ± mefedron skilde sig signifikant från respektive kontroller (p <0,0001 för alla behandlingar).
http://bbzzzsvqcrqtki6umym6itiixfhni37ybtt7mkbjyxn2pgllzxf2qgyd.onion/index.php?attachments/x0ik9hdwsw-jpg.4799/&hash=f31155ada4e59f22a35f9110aaa6a7e5
Fig. 4
Effekter av mefedron på metamfetamininducerad hypertermi. Möss behandlades med de angivna doserna av mefedron (MEPH) 30 minuter före varje injektion av 2,5 (a) eller 5,0 mg/kg (b) metamfetamin (METH). Kärntemperaturen mättes med 20 minuters intervall med telemetri med början 60 minuter före den första injektionen av metamfetamin. De 4 metamfetamininjektionerna indikeras av pilarna som vilar på x-axeln. Data uttrycks som genomsnittlig kroppstemperatur för 6-8 möss per grupp. SEM var alltid <10% av medelvärdet och utelämnas för tydlighetens skull.
Effekter av mefedron på amfetamin- och MDMA-inducerad neurotoxicitet
För att testa om de förstärkande effekterna av mefedron på metamfetamin kunde utsträckas till andra neurotoxiska amfetaminer, behandlades möss med denna β-ketoamfetamin (20 mg/kg) plus amfetamin (4X 5 mg/kg) eller MDMA (4X 20 mg/kg) och resultaten presenteras i figur 5. Kom ihåg att mefedron i sig inte minskar striatal DA, DAT eller TH (Angoa-Perez et al. 2012). Huvudeffekten av läkemedel (F5,27 = 27.18, p <0.0001) var mycket signifikant genom envägs ANOVA för DA-reduktioner (Fig. 5a). Det kan också ses i Fig. 5a att alla behandlingar med amfetamin (p <0.001) eller MDMA (p <0.001) ensam eller i kombination med mephedrone (p <0.0001 för båda läkemedlen) sänkte DA-nivåerna signifikant från kontroll. Mephedrone förbättrade signifikant DA-reduktioner orsakade av amfetamin (p <0.01) eller MDMA (p <0.01). Fig. 5b visar liknande effekter av kombinationsläkemedelsbehandlingar på DAT-nivåer i striatum. Huvudeffekten av läkemedel (F4,49 = 42.63, p <0.0001) var mycket signifikant av envägs ANOVA för DAT. Det kan också ses i fig. 5b att alla behandlingar med amfetamin eller MDMA var signifikant (p <0.0001 för alla) lägre jämfört med kontroll. Mephedrone förbättrade också signifikant DAT-reduktioner orsakade av antingen amfetamin eller MDMA (p <0.0001 i båda fallen). Slutligen, Fig. 5c visar att läkemedlets huvudeffekt (F4,50 = 75.06, p <0.0001) var mycket signifikant av envägs ANOVA för minskningar i TH. Det kan också ses i fig. 5c att alla behandlingar med amfetamin eller MDMA var signifikant (p < 0,0001 för alla) lägre i jämförelse med kontroll. Mephedrone förbättrade också signifikant TH-reduktioner orsakade av antingen amfetamin eller MDMA (p <0.0001 i båda fallen)
http://bbzzzsvqcrqtki6umym6itiixfhni37ybtt7mkbjyxn2pgllzxf2qgyd.onion/index.php?attachments/onvypkgxvn-jpg.4800/&hash=f31155ada4e59f22a35f9110aaa6a7e5
Fig. 5
Effekter av mefedron på amfetamin- eller MDMA-inducerad DA nervändande neurotoxicitet. Möss behandlades med 20 mg/kg mefedron (MEPH) 30 minuter före varje injektion av 5,0 mg/kg amfetamin (AMPH) eller 20 mg/kg MDMA och offrades 2d efter behandling för bestämning av striatala nivåer av (a) DA med HPLC. (b) DAT och (c) TH bestämdes genom immunoblotting och blottar kvantifierades med hjälp av ImageJ. Representativa immunoblots för DAT och TH ingår som insatser i paneler (b) respektive (c) och behandlingar för båda panelerna indikeras med 1,5: kontroll; 2,6: MEPH; 3: AMPH; 4: AMPH + MEPH; 7: MDMA; och 8: MDMA + MEPH. Data är medelvärde ± SEM för 5-12 möss i varje grupp. **p <0,001 eller ***p <0,0001 vs kontroll och #p <0,01 eller ###p <0,0001 vs AMPH eller MDMA (Tukey's multipla jämförelsetest).
Effekter av nomifensin på metamfetamininducerad neurotoxicitet
Nomifensine, en kraftfull DAT-blockerare utan känd missbruks- eller neurotoxisk potential, testades för sin förmåga att skydda mot metamfetamininducerad neurotoxicitet och för kontrast till mefedronets verkan på toxiciteten för DA-nervändningar orsakade av metamfetamin, amfetamin och MDMA. Resultaten i fig. 6a visar att läkemedlets huvudeffekt (F3,16 = 63,39, p < 0,0001) på DA-nivåer var mycket signifikant genom envägs ANOVA. Nomifensine ensam förändrade inte DA-nivåerna men minskningen orsakad av metamfetamin (p <0,0001) reverserades något men signifikant av nomifensine (p <0,01). Huvudeffekten av läkemedel (F3,20 = 16.78, p <0.0001) på DAT-nivåer var mycket signifikant av envägs ANOVA som visas i Fig. 6b. Nomifensine förändrade inte DAT-nivåer men gav signifikant skydd (p <0.001) mot minskningen av striatal DAT orsakad av metamfetamin (p <0.0001) jämfört med kontroll. Slutligen, Fig. 6c visar att läkemedlets huvudeffekt (F3,15 = 14.10, p <0.0001) på TH-nivåer var mycket signifikant av envägs ANOVA. Som sett för DA och DAT, minskningen av TH orsakad av metamfetamin (p <0.0001) förhindrades något men signifikant av nomifensin (p <0.01).
http://bbzzzsvqcrqtki6umym6itiixfhni37ybtt7mkbjyxn2pgllzxf2qgyd.onion/index.php?attachments/daczg9cwfv-jpg.4801/&hash=f31155ada4e59f22a35f9110aaa6a7e5
Fig. 6
Effekter av nomifensin på metamfetamininducerad DA nervändande neurotoxicitet. Möss behandlades med 5,0 mg/kg nomifensin (NOM) 30 minuter före varje injektion av 5,0 mg/kg metamfetamin (METH) och offrades 2 dagar senare för bestämning av striatala nivåer av (a) DA med HPLC. (b) DAT och (c) TH bestämdes genom immunoblotting och blottar kvantifierades med hjälp av ImageJ. Representativa immunoblots för DAT och TH ingår som insatser i panelerna (b) och (c) respektive. Data är medelvärde plus SEM för 5-7 möss per grupp. ***p <0,0001 vs kontroll (C) och #p <0,01 eller ##p <0,001 vs enbart metamfetamin (Tukeys multipla jämförelsetest).
Diskussion
Syftet med den aktuella studien var att avgöra om mefedron skulle förhindra DA-nervavslutande toxicitet orsakad av metamfetamin. Baserat på dess kemiska likhet med metamfetamin och MDMA förväntades det ursprungligen att mefedron skulle utöva skadliga effekter på DA-neuronsystemet. Flera studier visade dock nästan samtidigt att mefedron inte var giftigt för DA-nervändarna (Angoa-Perez et al. 2012, Baumann et al. 2012, Hadlock et al. 2011). Frågan om huruvida detta läkemedel orsakar skador på 5-HT-neuronsystemet förblir öppen. En studie rapporterade ihållande minskningar av 5-HT-nervändfunktion (Hadlock et al. 2011) medan en annan fann att mefedron inte orsakade skada (Baumann et al. 2012). Mefedron interagerar med DA-nervänden på ett sätt som tyder på att det verkligen stimulerar frisättning och blockerar DA-återupptag via dess interaktioner med DAT. En viktig aspekt av metamfetamins neurotoxiska verkningsmekanism är dess förmåga att få tillgång till DA-nervändarna genom DAT och störa DA-homeostasen (Sulzer 2011). Om detta tidiga steg i metamfetaminets neurotoxiska kaskad förhindras genom hämning av DAT, förhindras toxicitet (Pu et al. 1994, Poth et al. 2012, Marek et al. 1990, Schmidt och Gibb 1985). Vi antog att mefedron skulle kunna ha samma skyddande egenskaper som andra DAT-hämmare, men observerade istället en signifikant ökning av toxiciteten. Denna interaktion sågs med hjälp av två olika doser av metamfetamin som orsakar måttlig eller allvarlig skada på DA-nervändarna (4X 2,5 respektive 5,0 mg/kg). Denna förstärkande effekt av mefedron var inte begränsad till metamfetamin utan utsträcktes till amfetamin och MDMA, två droger som ofta sammissbrukas med mefedron och andra β-ketoamfetaminer (Feyissa och Kelly 2008, Schifano et al. 2011, Kelly 2011). Trots att mefedron inte orsakar toxicitet för åtminstone DA-nervändarna i striatum förstärker det därför de neurotoxiska effekterna av andra missbrukssubstanser. Detta nya fynd bör ge mefedronmissbruk en ännu skarpare bild, eftersom dess brist på inneboende neurotoxicitet kan få det att framstå som ofarligt.
Hypertermi är en ofta rapporterad akut biverkning av intag av metamfetamin (Greene et al. 2008) och β-ketoamfetamin hos människor (Borek och Holstege 2012, Prosser och Nelson 2012). Liksom metamfetamin orsakar många av β-ketoamfetaminläkemedlen också betydande höjningar av kärntemperaturen hos gnagare (Angoa-Perez m.fl. 2012, Hadlock m.fl. 2011, Baumann m.fl. 2012, Rockhold m.fl. 1997). Även om den hypertermi som orsakas av metamfetamin kan bidra till dess morfologiska och neuronala skadeverkningar, är det inte nödvändigtvis så att hypertermi är den direkta orsaken till dessa effekter (Kiyatkin och Sharma 2009). Vi registrerade kroppstemperaturen hos möss som behandlats med mefedron och metamfetamin och observerade att den kombinerade behandlingen inte ökade temperaturen utöver de maximala ökningar som sågs efter endera läkemedlet ensamt. Metamfetamin orsakade en dosrelaterad ökning av kroppstemperaturen och denna hypertermi var oföränderlig över hela det testade dosintervallet för mefedron. Faktum är att det fall i kroppstemperatur efter injektion som observerades efter mefedronbehandling (Angoa-Perez et al. 2012) bibehölls vid högre doser av mefedron plus metamfetamin. Även om den läkemedelsinducerade hypertermi inte förstärktes av kombinerad läkemedelsbehandling, var de neurotoxiska effekterna additiva. Därför förefaller det, åtminstone i det aktuella fallet, som om de neurotoxiska effekterna av metamfetamin kan förstärkas av mefedron på ett sätt som är oberoende av hypertermi.
Mefedron hämmar tydligt DAT-funktionen och blockerar återupptaget av DA in vitro (Lopez-Arnau et al. 2012, Hadlock et al. 2011, Kehr et al. 2011, Martinez-Clemente et al. 2012, Cozzi et al. 1999). Mefedron undantränger WIN-35 428 från dess bindningsställe på DAT, vilket tyder på att det är en konkurrenskraftig hämmare av DA-upptag (Martinez-Clemente et al. 2012, Lopez-Arnau et al. 2012). Mefedronets styrka i detta avseende är mycket lik metamfetaminets (Cozzi et al. 1999) och MDMA:s (Escubedo et al. 2011). Det är inte känt om mefedron transporteras av DAT, men det gör metkatinon (Cozzi och Foley 2003). Nomifensin och amfonsyra, som binder till DAT och hämmar DA-upptaget, ger ett betydande skydd mot metamfetamininducerad neurotoxicitet (Pu et al. 1994, Marek et al. 1990, Schmidt och Gibb 1985, Poth et al. 2012) och möss som saknar DAT är resistenta mot metamfetaminets neuronala toxicitet (Fumagalli et al. 1998). Att veta att mefedron är icke-neurotoxiskt och en DAT-blockerare leder till förutsägelsen att det borde förhindra toxicitet. Vi testade nomifensine i detta avseende som en positiv kontroll och bekräftade att det skyddar mot metamfetamininducerad utarmning av DA, DAT och TH. Nomifensine hämmar också norepinefrintransportören (Brogden et al. 1979) men denna egenskap kan inte förklara de nuvarande resultaten eftersom de flesta β-ketoamfetaminer inklusive mefedron hämmar norepinefrintransportören och blockerar norepinefrinupptag (Kelly 2011, Rothman et al. 2003, Cozzi et al. 1999, Sogawa et al. 2011, Lopez-Arnau et al. 2012). En roll för det neuronala 5-HT-systemet i vissa av mefedronets farmakologiska verkningar är möjlig mot bakgrund av detta läkemedels förmåga att, liksom MDMA (Yamamoto et al. 1995), orsaka utflöde av striatal DA via dess interaktioner med 5-HT2A-receptorer (Lopez-Arnau et al. 2012, Martinez-Clemente et al. 2012). Den hyperlokomotion som orsakas av mefedron är beroende av endogent 5-HT (Lopez-Arnau et al. 2012) och detta läkemedel stimulerar också frisättningen av 5-HT och hämmar dess upptag in vitro (Sogawa et al. 2011, Cozzi et al. 1999, Nagai et al. 2007, Hadlock et al. 2011, Lopez-Arnau et al. 2012, Martinez-Clemente et al. 2012) och in vivo (Baumann et al. 2012, Kehr et al. 2011). Vi kan dock utesluta en roll för endogent 5-HT i DA-neurotoxiciteten hos åtminstone metamfetamin genom att visa att möss som är genetiskt utarmade på 5-HT behåller sin känslighet för neurotoxicitet (Thomas et al. 2010).
Mefedron kan förstärka metamfetamins neurotoxicitet genom flera möjliga mekanismer. För det första kan mefedron interagera med VMAT för att orsaka läckage av DA till cytoplasman i den presynaptiska nervändan. Behandlingar som ökar den cytoplasmatiska poolen (dvs. läkemedelsfrisättande) av DA ökar metamfetaminets neurotoxicitet (Thomas et al. 2008, Thomas et al. 2009, Schmidt et al. 1985). Denna mekanism är inte sannolik eftersom metkatinon endast interagerar svagt med VMAT (Cozzi et al. 1999). För det andra kan kombinationen av mefedron plus metamfetamin ha en synergistisk effekt på icke-vesikulär frisättning av DA, men denna möjlighet verkar också osannolik mot bakgrund av resultat som visar att behandling av DAT- eller SERT-uttryckande CHO-celler med metylon plus metamfetamin inte har en additiv effekt på DA- eller 5-HT-frisättning (Sogawa et al. 2011). För det tredje kan mefedron interagera med DAT på ett nytt sätt som bidrar till additiv toxicitet. Det har visats att metylon i kombination med metamfetamin orsakar synergistisk cytotoxicitet i CHO-celler som uttrycker DAT eller SERT men inte i CHO-celler av vildtyp som saknar transportörer (Sogawa et al. 2011). Den cytotoxicitet som ses i odlade celler i dessa studier (dvs. LDH-frisättning) skiljer sig mycket från skadorna på DA-nervändarna som orsakas av metamfetamin, men denna mekanism antyder en intressant men odefinierad roll för DAT i ökad cytotoxicitet. Slutligen kan mefedron förändra metamfetaminets metabolism. Mefedron metaboliseras främst genom N-demetylering (Meyer och Maurer 2010), liksom metamfetamin och MDMA (Caldwell 1976). Stöd för denna mekanism kommer från demonstrationen att metamfetamin och MDMA ömsesidigt hämmar produktionen av sina respektive primära metaboliter och höjer plasmanivåerna av läkemedlen över dem som ses efter administrering av endera läkemedlet ensamt (Kuwayama et al. 2012). De doser av mefedron som används för närvarande och i vår tidigare studie (Angoa-Perez et al. 2012) är visserligen höga men inte neurotoxiska och ligger inom det intervall som missbrukas av människor (McErath och O'Neill 2011). Därför kan mefedron fungera som MDMA för att öka plasmanivåerna av metamfetamin genom att hämma dess metabolism. En fördjupad farmakokinetisk analys kommer att krävas för att bekräfta denna senare möjlighet.
Missbruket av β-ketoamfetaminer ökar i en alarmerande takt och mefedron är nu en av de vanligaste drogerna efter cannabis, MDMA och kokain (Morris 2010, Winstock et al. 2011b). Dessutom framkallar mefedron starkare känslor av begär hos människor jämfört med MDMA (Brunt m.fl. 2011) och användare som snortar mefedron anser att det är mer beroendeframkallande än kokain (Winstock m.fl. 2011b). Mefedron konsumeras av människor på ett binge-liknande sätt (dvs. "stacking") och tas ofta tillsammans med andra droger som cannabis och amfetaminpsykostimulantia (Schifano m.fl. 2011, Fass m.fl. 2012, Winstock m.fl. 2011a, Kelly 2011, Torrance och Cooper 2010). Mefedron förekommer allt oftare i tabletter som säljs som MDMA (Brunt m.fl. 2011) och användningen av mefedron kommer sannolikt att överstiga användningen av MDMA i takt med att renheten hos den senare drogen fortsätter att sjunka (Brunt m.fl. 2011, Tanner-Smith 2006, Teng m.fl. 2006). Baserat på de vanliga missbruksmönstren för mefedron och andra ingredienser i "badsalt" är det viktigt att överväga om ytterligare hälsorisker uppstår hos människor när dessa droger avsiktligt eller oavsiktligt kombineras med amfetamin. Våra resultat som visar att åtminstone mefedron avsevärt förstärker neurotoxiciteten för DA-nervändarna i striatum som orsakas av metamfetamin, amfetamin och MDMA avslöjar en särskilt farlig och oväntad egenskap hos denna β-ketoamfetamin.
Förkortningar som används
5-HT serotonin
DA dopamin
DAT DA-transportör
MDMA 3,4-metylendioximetamfetamin
TH tyrosinhydroxylas
VMAT vesikulär monoamintransportör